

ORE9, an F-box protein that regulates leaf senescence in Arabidopsis
- PMID: 11487692
- PMCID: PMC139127
- DOI: 10.1105/tpc.010061
ORE9, an F-box protein that regulates leaf senescence in Arabidopsis
Abstract
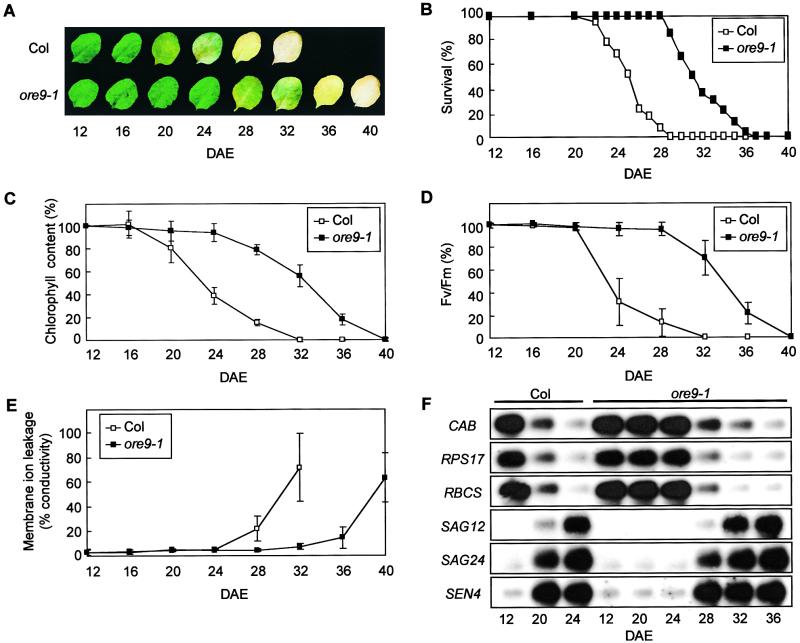
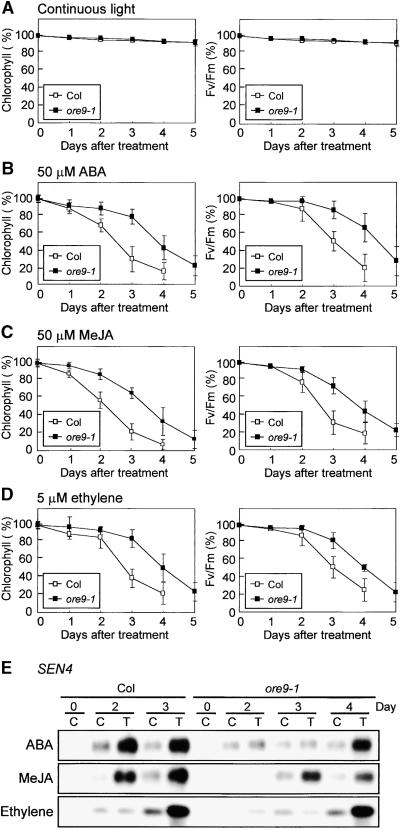
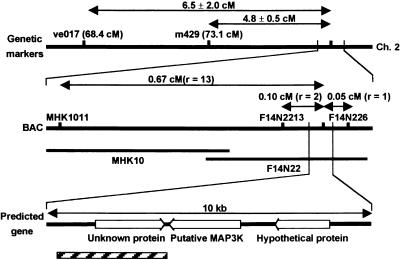
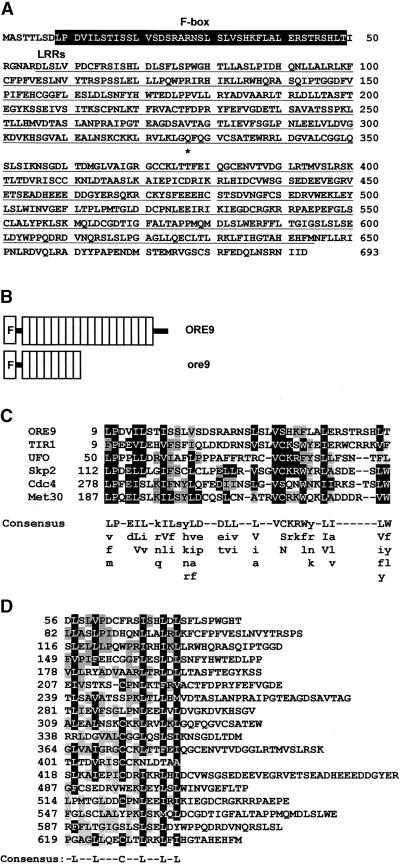
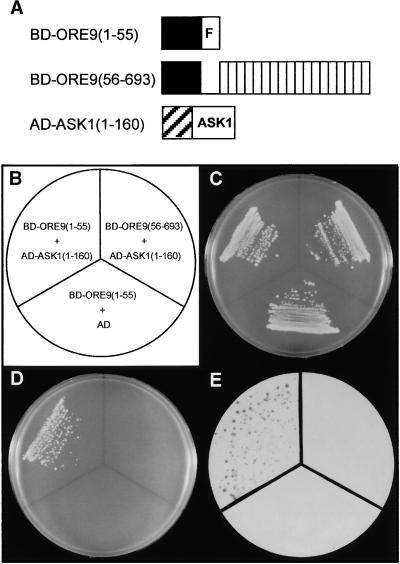
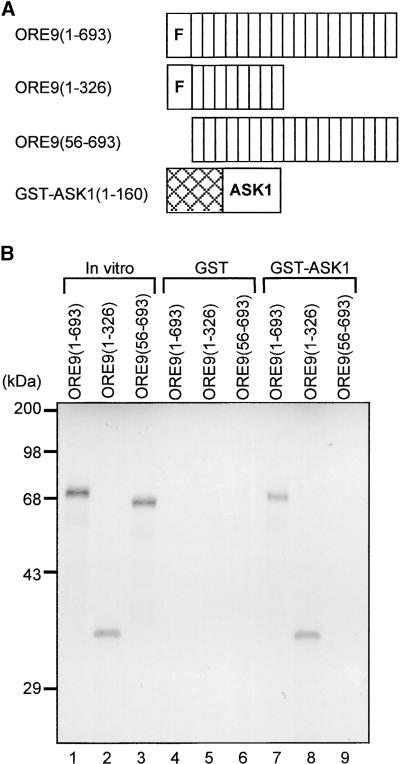
Senescence is a sequence of biochemical and physiological events that constitute the final stage of development. The identification of genes that alter senescence has practical value and is helpful in revealing pathways that influence senescence. However, the genetic mechanisms of senescence are largely unknown. The leaf of the oresara9 (ore9) mutant of Arabidopsis exhibits increased longevity during age-dependent natural senescence by delaying the onset of various senescence symptoms. It also displays delayed senescence symptoms during hormone-modulated senescence. Map-based cloning of ORE9 identified a 693-amino acid polypeptide containing an F-box motif and 18 leucine-rich repeats. The F-box motif of ORE9 interacts with ASK1 (Arabidopsis Skp1-like 1), a component of the plant SCF complex. These results suggest that ORE9 functions to limit leaf longevity by removing, through ubiquitin-dependent proteolysis, target proteins that are required to delay the leaf senescence program in Arabidopsis.
Figures
References
-
- Aharoni, N. (1989). Interrelationship between ethylene and growth regulators in senescence of lettuce leaf disk. J. Plant Growth Regul. 8, 309–317.
-
- Bai, C., Sen, P., Hofmann, K., Ma, L., Goebl, M., Harper, J.W., and Elledge, S.J. (1996). SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box. Cell 86, 263–274. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
