

EMF1, a novel protein involved in the control of shoot architecture and flowering in Arabidopsis
- PMID: 11487698
- PMCID: PMC139134
- DOI: 10.1105/tpc.010094
EMF1, a novel protein involved in the control of shoot architecture and flowering in Arabidopsis
Abstract
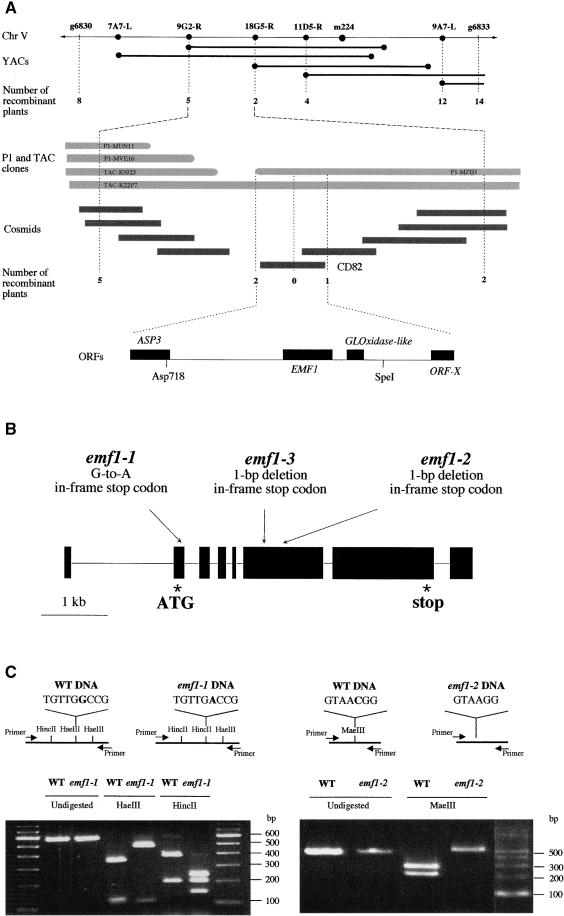
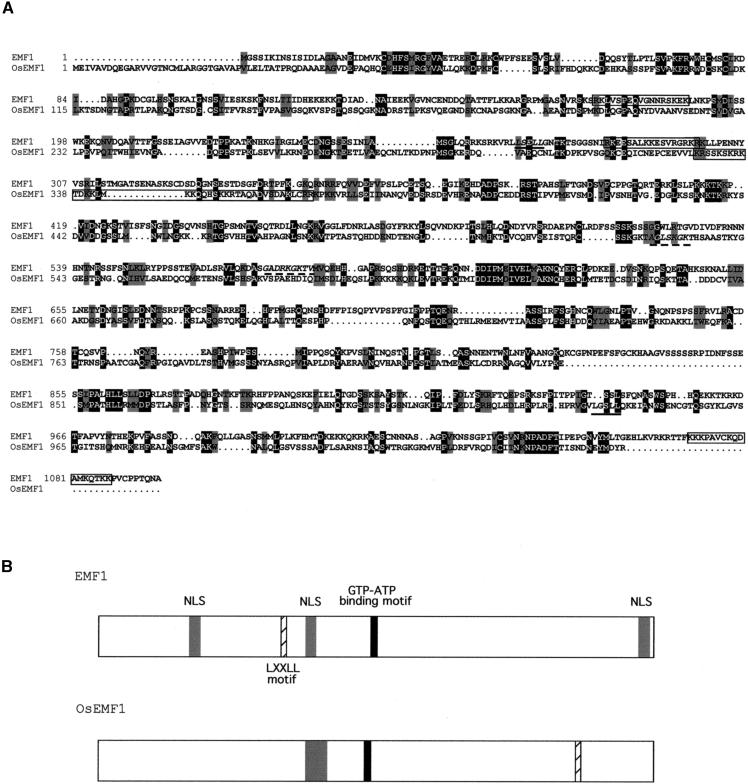
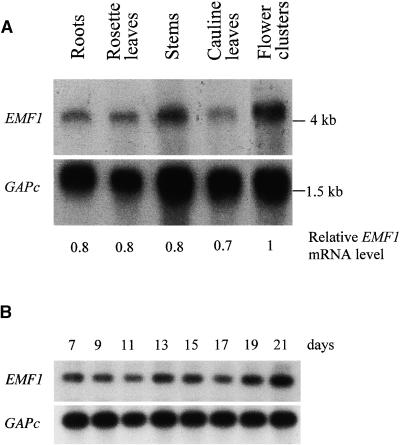
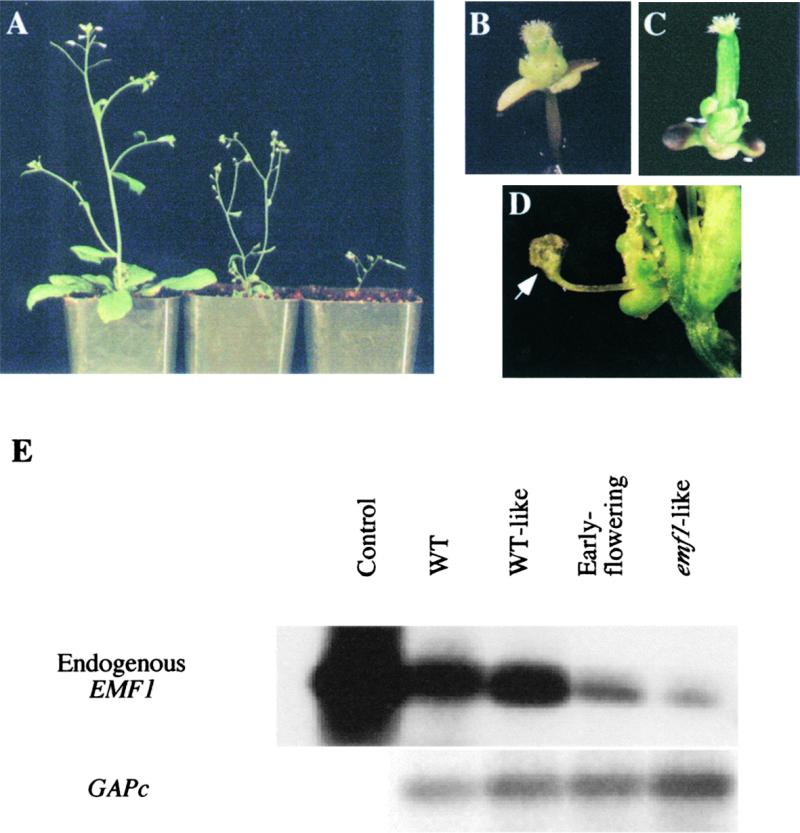
Shoot architecture and flowering time in angiosperms depend on the balanced expression of a large number of flowering time and flower meristem identity genes. Loss-of-function mutations in the Arabidopsis EMBRYONIC FLOWER (EMF) genes cause Arabidopsis to eliminate rosette shoot growth and transform the apical meristem from indeterminate to determinate growth by producing a single terminal flower on all nodes. We have identified the EMF1 gene by positional cloning. The deduced polypeptide has no homology with any protein of known function except a putative protein in the rice genome with which EMF1 shares common motifs that include nuclear localization signals, P-loop, and LXXLL elements. Alteration of EMF1 expression in transgenic plants caused progressive changes in flowering time, shoot determinacy, and inflorescence architecture. EMF1 and its related sequence may belong to a new class of proteins that function as transcriptional regulators of phase transition during shoot development.
Figures
References
-
- Alvarez, J., Guli, C.L., Yu, X.-H., and Smyth, D.R. (1992). terminal flower: A gene affecting inflorescence development in Arabidopsis thaliana. Plant J. 2, 103–116.
-
- Bechtold, N., Ellis, J., and Pelletier, G. (1993). In planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C. R. Acad. Sci. Paris Life Sci. 316, 1194–1199.
-
- Blazquez, M. (2000). Flower development pathways. J. Cell Sci. 113, 3547–3548. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
