

Activation of salicylic acid-induced protein kinase, a mitogen-activated protein kinase, induces multiple defense responses in tobacco
- PMID: 11487699
- PMCID: PMC139126
- DOI: 10.1105/tpc.010044
Activation of salicylic acid-induced protein kinase, a mitogen-activated protein kinase, induces multiple defense responses in tobacco
Abstract
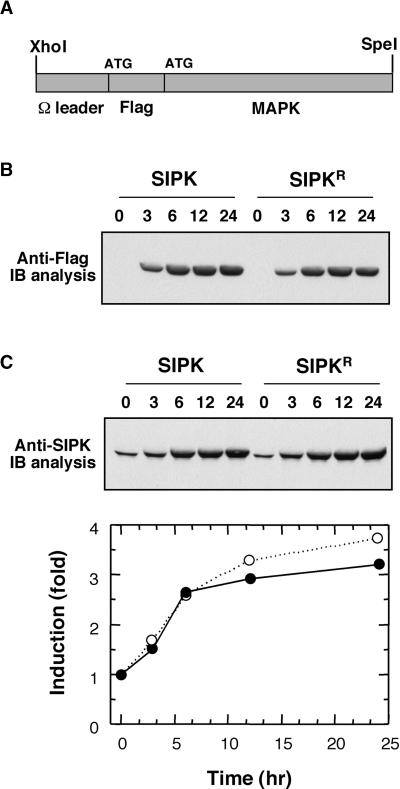
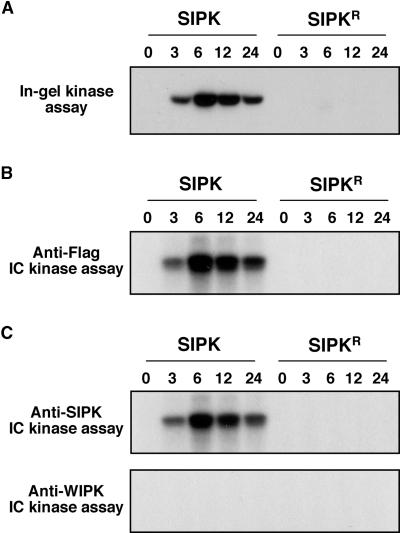
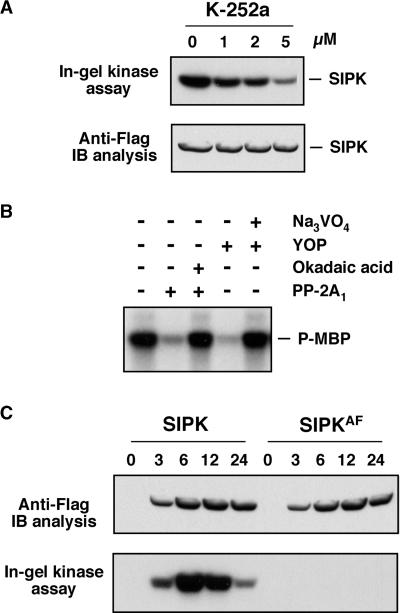
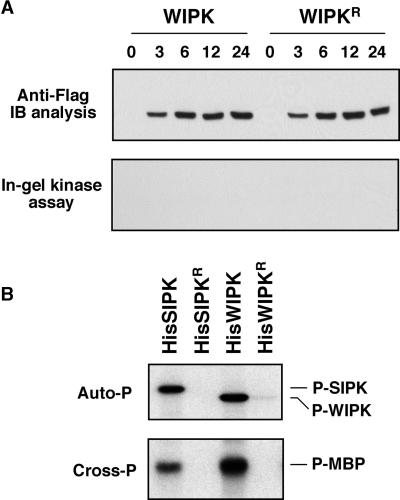
The activation of mitogen-activated protein kinases (MAPKs) is one of the earliest responses in plants challenged by avirulent pathogens or cells treated with pathogen-derived elicitors. Expression of a constitutively active MAPK kinase, NtMEK2(DD), in tobacco induces the expression of defense genes and hypersensitive response-like cell death, which are preceded by the activation of two endogenous MAPKs, salicylic acid-induced protein kinase (SIPK) and wounding-induced protein kinase (WIPK). However, the roles that SIPK and WIPK each play in the process are unknown. Here we report that SIPK alone is sufficient to activate these defense responses. In tobacco leaves transiently transformed with SIPK under the control of a steroid-inducible promoter, the induction of SIPK expression after the application of dexamethasone, a steroid, leads to an increase of SIPK activity. The increase of SIPK activity is dependent on the phosphorylation of newly synthesized SIPK by its endogenous upstream kinase. In contrast, the expression of WIPK under the same conditions fails to increase its activity, even though the protein accumulates to a similar level. Studies using chimeras of SIPK and WIPK demonstrated that the C terminus of SIPK contains the molecular determinant for its activation, which is rather surprising because the N termini of SIPK and WIPK are more divergent. SIPK has been implicated previously in the regulation of both plant defense gene activation and hypersensitive response-like cell death based on evidence from pharmacological studies using kinase inhibitors. This gain-of-function study provided more direct evidence for its role in the signaling of multiple defense responses in tobacco.
Figures
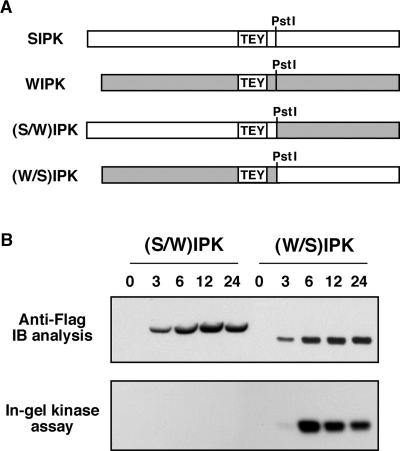
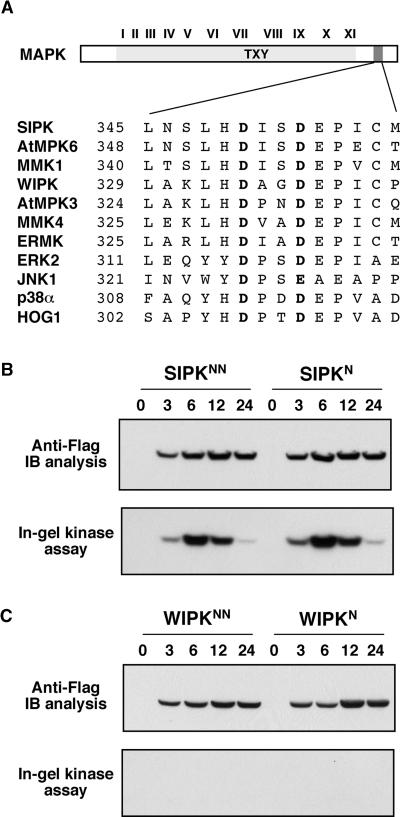

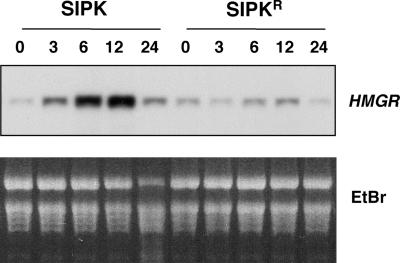
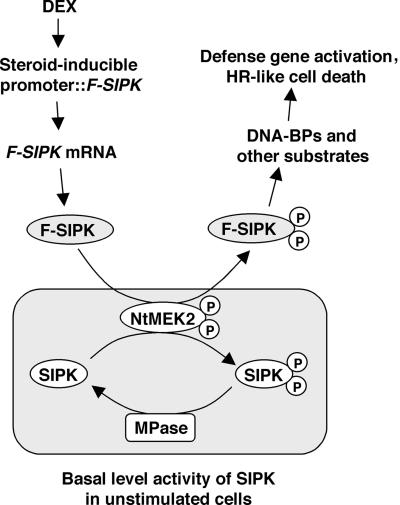
References
-
- Aoyama, T., and Chua, N.-H. (1997). A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 11, 605–612. - PubMed
-
- Arabidopsis Genome Initiative. (2000). Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 108, 796–815. - PubMed
-
- Brunner, D., Oellers, N., Szabad, J., Biggs, W.H., III, Zipursky, S.L., and Hafen, E. (1994). A gain-of-function mutation in Drosophila MAP kinase activates multiple receptor tyrosine kinase signaling pathways. Cell 76, 875–888. - PubMed
-
- Caffrey, D.R., O'Neill, L.A.J., and Shields, D.C. (1999). The evolution of the MAP kinase pathways: Coduplication of interacting proteins leads to new signaling cascades. J. Mol. Evol. 49, 567–582. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
