

LcrG-LcrV interaction is required for control of Yops secretion in Yersinia pestis
- PMID: 11489861
- PMCID: PMC95384
- DOI: 10.1128/JB.183.17.5082-5091.2001
LcrG-LcrV interaction is required for control of Yops secretion in Yersinia pestis
Abstract
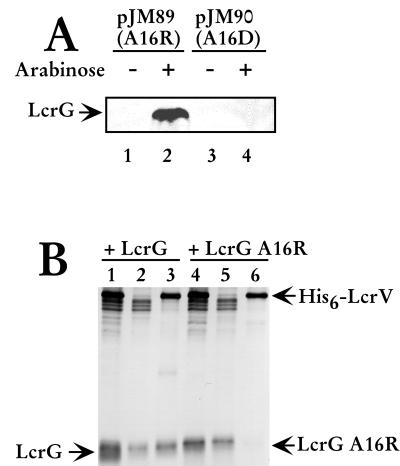
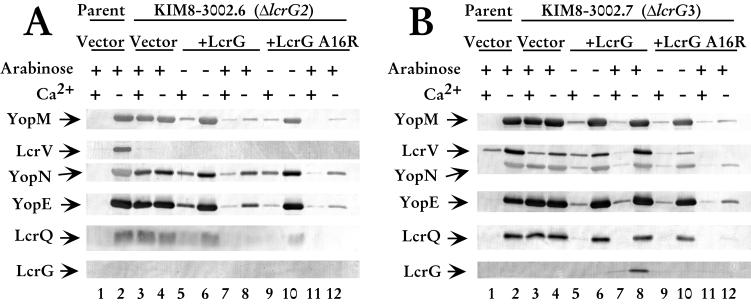
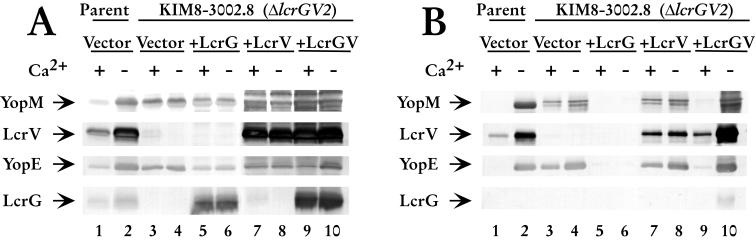
Yersinia pestis expresses a set of plasmid-encoded virulence proteins called Yops and LcrV that are secreted and translocated into eukaryotic cells by a type III secretion system. LcrV is a multifunctional protein with antihost and positive regulatory effects on Yops secretion that forms a stable complex with a negative regulatory protein, LcrG. LcrG has been proposed to block the secretion apparatus (Ysc) from the cytoplasmic face of the inner membrane under nonpermissive conditions for Yops secretion, when levels of LcrV in the cell are low. A model has been proposed to describe secretion control based on the relative levels of LcrG and LcrV in the bacterial cytoplasm. This model proposes that under secretion-permissive conditions, levels of LcrV are increased relative to levels of LcrG, so that the excess LcrV titrates LcrG away from the Ysc, allowing secretion of Yops to occur. To further test this model, a mutant LcrG protein that could no longer interact with LcrV was created. Expression of this LcrG variant blocked secretion of Yops and LcrV under secretion permissive conditions in vitro and in a tissue culture model. These results agree with the previously described secretion-blocking activity of LcrG and demonstrate that the interaction of LcrV with LcrG is necessary for controlling Yops secretion.
Figures

References
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons; 1998.
-
- Boyd A P, Sory M-P, Iriarte M, Cornelis G R. Heparin interferes with translocation of Yop proteins into HeLa cells and binds to LcrG, a regulatory component of the Yersinia Yop apparatus. Mol Microbiol. 1998;27:425–436. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
