

p70S6 kinase signals cell survival as well as growth, inactivating the pro-apoptotic molecule BAD
- PMID: 11493700
- PMCID: PMC55509
- DOI: 10.1073/pnas.171301998
p70S6 kinase signals cell survival as well as growth, inactivating the pro-apoptotic molecule BAD
Abstract
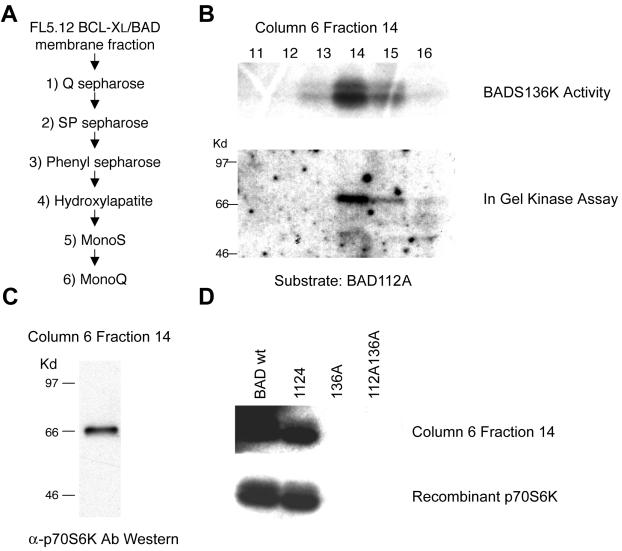
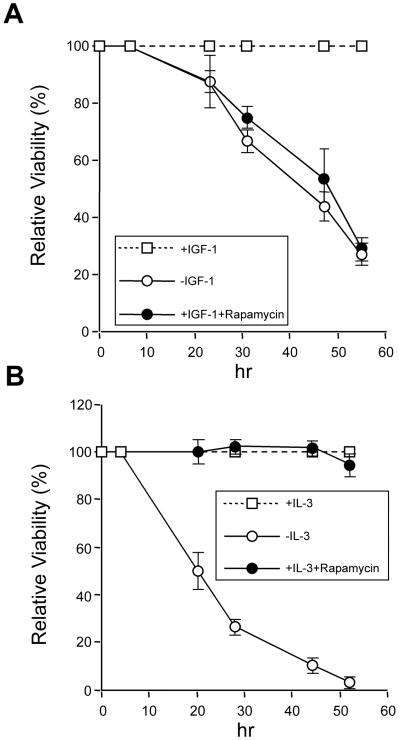
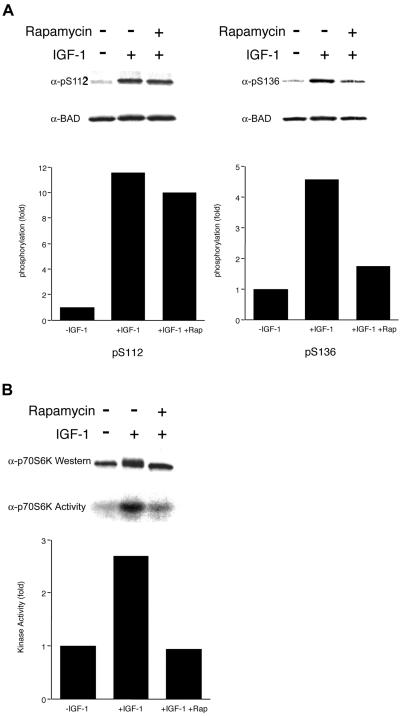
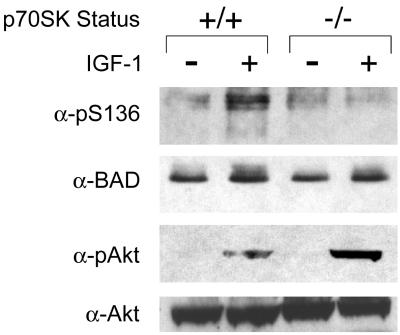
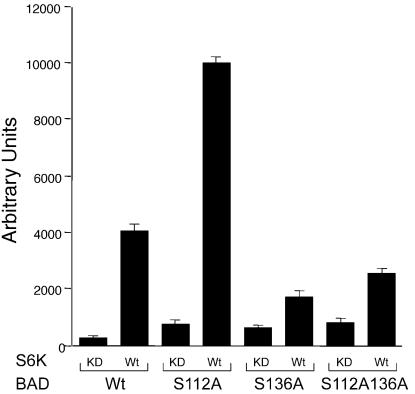
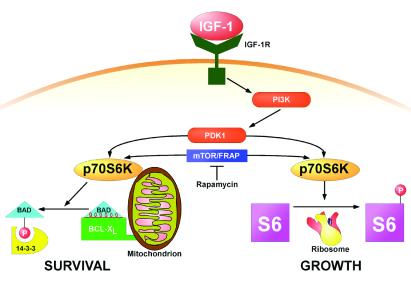
Cytokines often deliver simultaneous, yet distinct, cell growth and cell survival signals. The 70-kDa ribosomal protein S6 kinase (p70S6K) is known to regulate cell growth by inducing protein synthesis components. We purified membrane-based p70S6K as a kinase responsible for site-specific phosphorylation of BAD, which inactivates this proapoptotic molecule. Rapamycin inhibited mitochondrial-based p70S6K, which prevented phosphorylation of Ser-136 on BAD and blocked cell survival induced by insulin-like growth factor 1 (IGF-1). Moreover, IGF-1-induced phosphorylation of BAD Ser-136 was abolished in p70S6K-deficient cells. Thus, p70S6K is itself a dual pathway kinase, signaling cell survival as well as growth through differential substrates which include mitochondrial BAD and the ribosomal subunit S6, respectively.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
