

FANCC interacts with Hsp70 to protect hematopoietic cells from IFN-gamma/TNF-alpha-mediated cytotoxicity
- PMID: 11500375
- PMCID: PMC125562
- DOI: 10.1093/emboj/20.16.4478
FANCC interacts with Hsp70 to protect hematopoietic cells from IFN-gamma/TNF-alpha-mediated cytotoxicity
Abstract
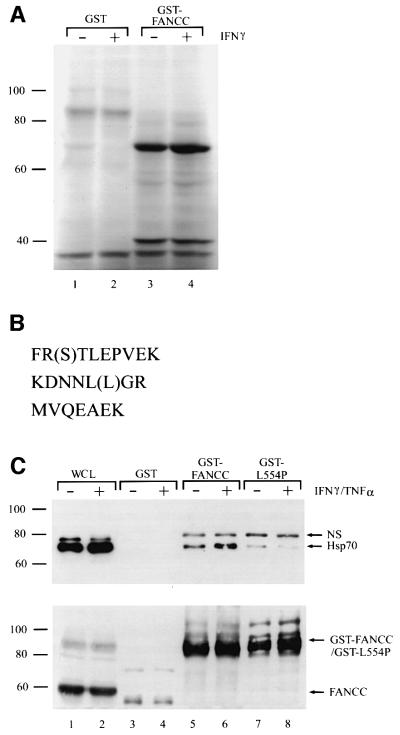
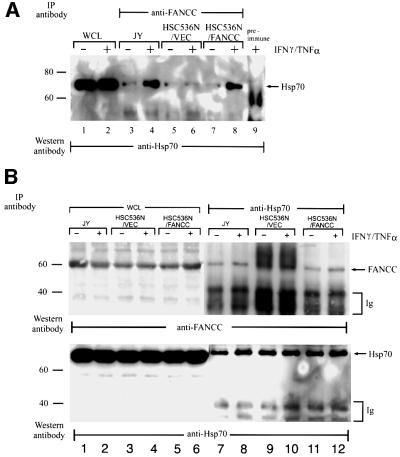
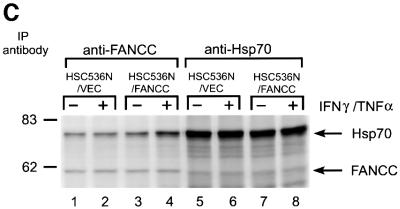
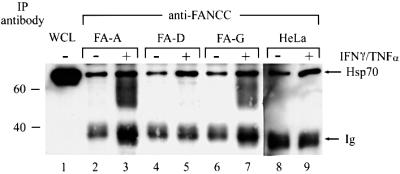
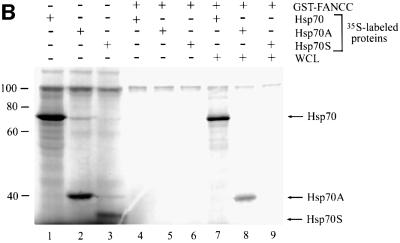
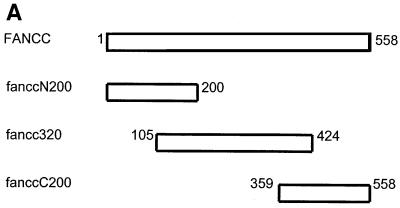
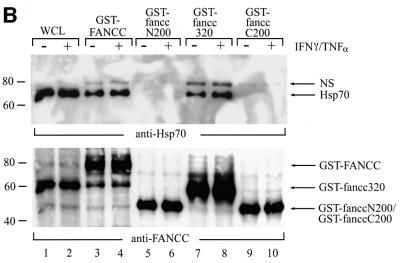
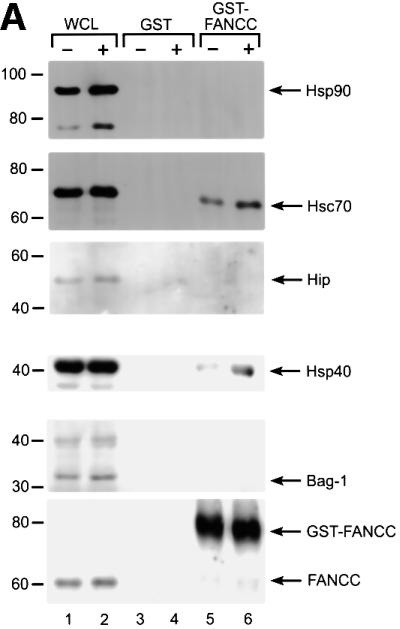
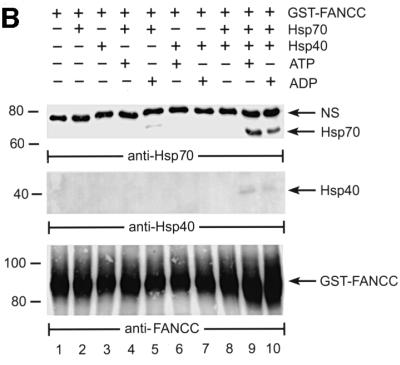
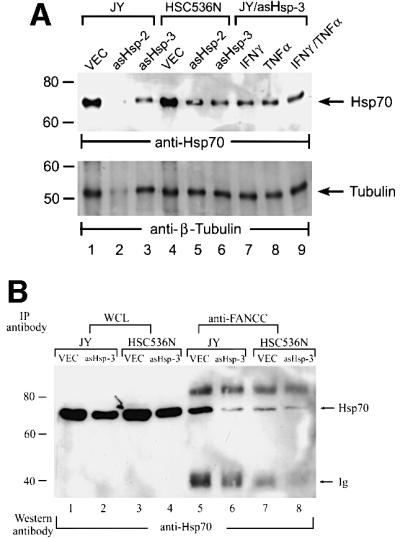
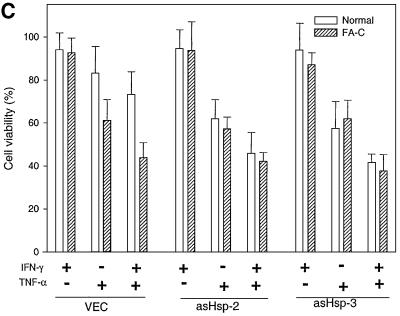
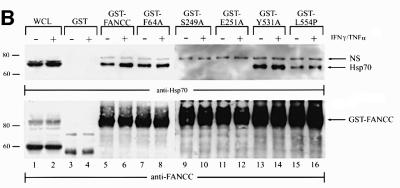
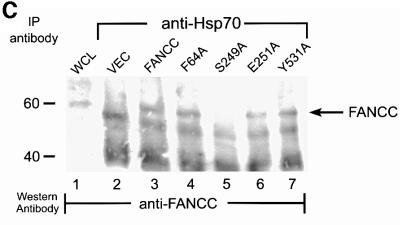
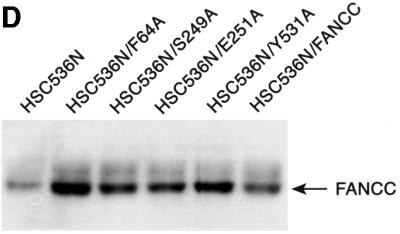
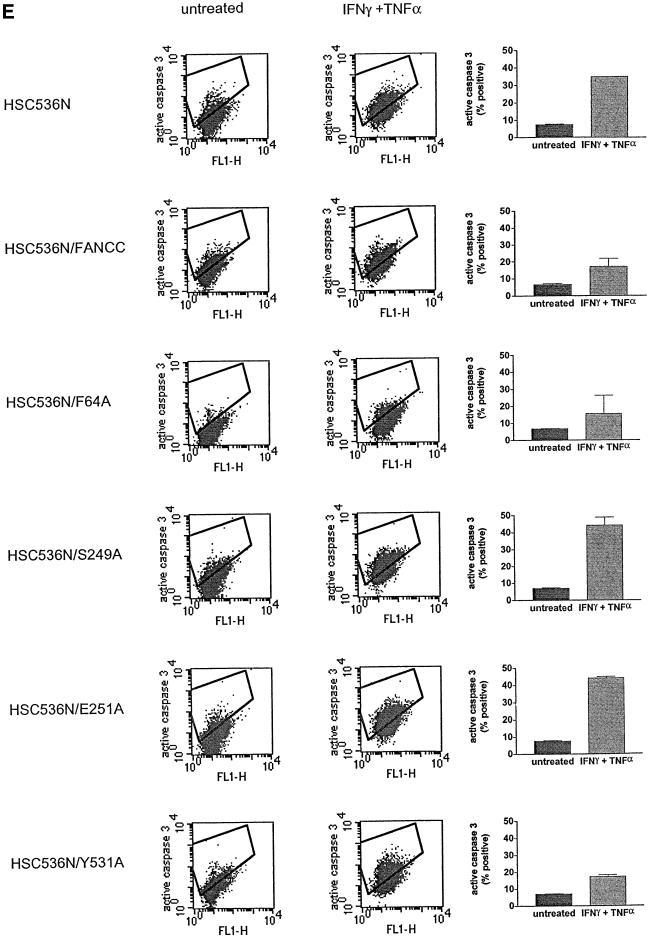
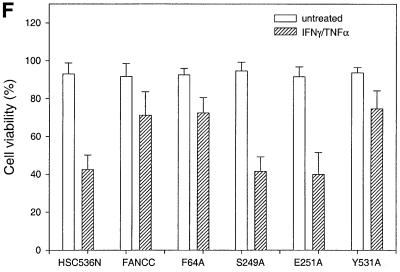
The Fanconi anemia (FA) complementation group C gene product (FANCC) functions to protect hematopoietic cells from cytotoxicity induced by interferon-gamma (IFN-gamma), tumor necrosis factor-alpha (TNF-alpha) and double-stranded RNA (dsRNA). Because apoptotic responses of mutant FA-C cells involve activation of interferon-inducible, dsRNA-dependent protein kinase PKR, we sought to identify FANCC-binding cofactors that may modulate PKR activation. We identified the molecular chaperone Hsp70 as an interacting partner of FANCC in lymphoblasts and HeLa cells using 'pull-down' and co-immunoprecipitation experiments. In vitro binding assays showed that the association of FANCC and Hsp70 involves the ATPase domain of Hsp70 and the central 320 residues of FANCC, and that both Hsp40 and ATP/ADP are required. In whole cells, Hsp70-FANCC binding and protection from IFN-gamma/TNF-alpha-induced cytotoxicity were blocked by alanine mutations located in a conserved motif within the Hsp70-interacting domain of FANCC. We therefore conclude that FANCC acts in concert with Hsp70 to prevent apoptosis in hematopoietic cells exposed to IFN-gamma and TNF-alpha.
Figures


References
-
- Agostini I., Popov,S., Li,J., Dubrovsky,L., Hao,T. and Bukrinsky,M. (2000) Heat-shock protein 70 can replace viral protein R of HIV-1 during nuclear import of the viral preintegration complex. Exp. Cell Res., 259, 398–403. - PubMed
-
- Auerbach A.D. and Allen,R.G. (1991) Leukemia and preleukemia in Fanconi anemia patients. A review of the literature and report of the International Fanconi Anemia Registry. Cancer Genet. Cytogenet., 51, 1–12. - PubMed
-
- Bohen S.P., Kralli,A. and Yamamoto,K.R. (1995) Hold ‘em and fold ‘em: chaperones and signal transduction. Science, 268, 1362–1365. - PubMed
-
- Buchwald M. and Moustacchi,E. (1998) Is Fanconi anemia caused by a defect in the processing of DNA damage? Mutat. Res., 408, 75–90. - PubMed
-
- Chappell T.G., Konforti,B.B., Schmid,S.L. and Rothman,J.E. (1987) The ATPase core of a clathrin uncoating protein. J. Biol. Chem., 262, 746–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
