

Overexpression of KNAT1 in lettuce shifts leaf determinate growth to a shoot-like indeterminate growth associated with an accumulation of isopentenyl-type cytokinins
- PMID: 11500537
- PMCID: PMC117138
- DOI: 10.1104/pp.126.4.1370
Overexpression of KNAT1 in lettuce shifts leaf determinate growth to a shoot-like indeterminate growth associated with an accumulation of isopentenyl-type cytokinins
Abstract
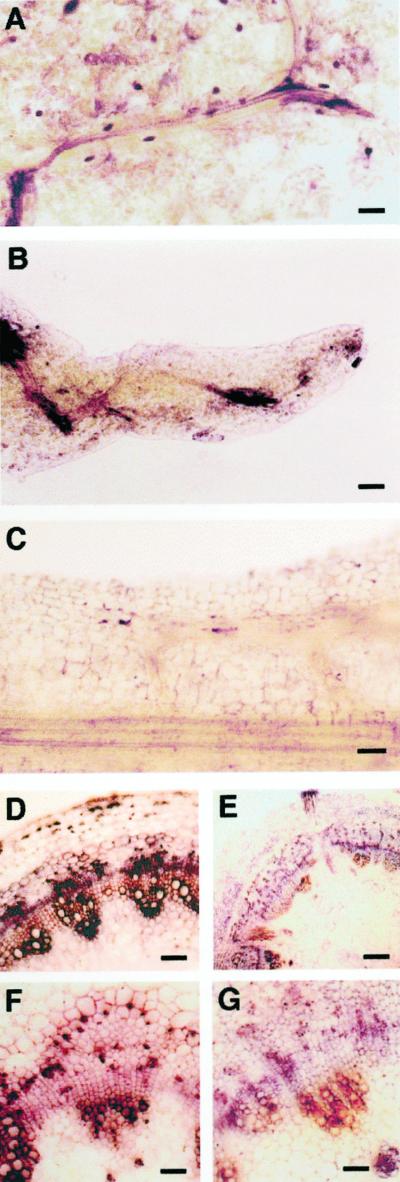
Leaves are specialized organs characterized by defined developmental destiny and determinate growth. The overexpression of Knotted1-like homeobox genes in different species has been shown to alter leaf shape and development, but a definite role for this class of genes remains to be established. Transgenics that overexpress Knotted1-like genes present some traits that are characteristic of altered cytokinin physiology. Here we show that lettuce (Lactuca sativa) leaves that overexpress KNAT1, an Arabidopsis kn1-like gene, acquire characteristics of indeterminate growth typical of the shoot and that this cell fate change is associated with the accumulation of specific types of cytokinins. The possibility that the phenotypic effects of KNAT1 overexpression may arise primarily from the modulation of local ratios of different cytokinins is discussed.
Figures


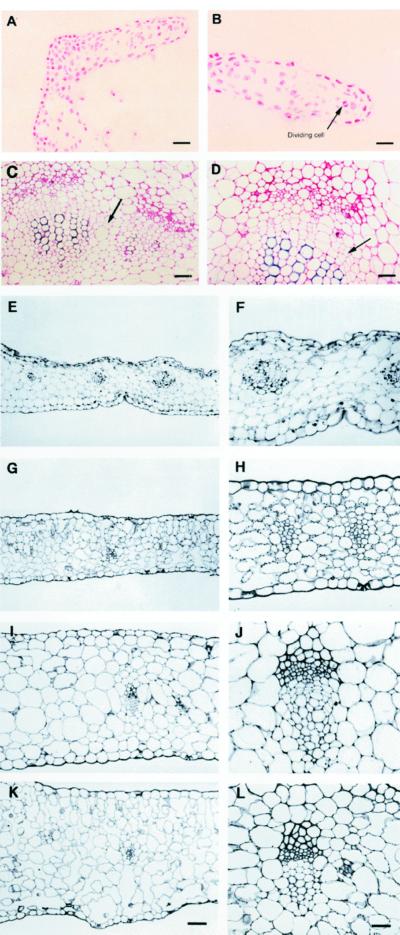
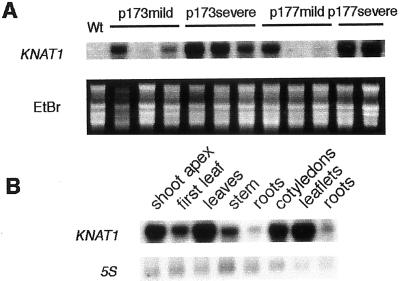
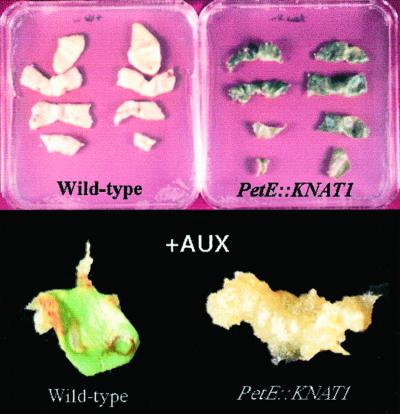
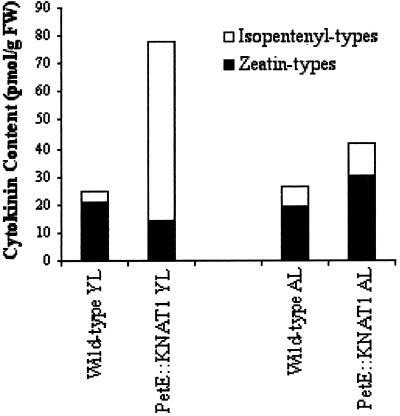
References
-
- Chan RL, Gago GM, Palena CM, Gonzalez DH. Homeoboxes in plant development. Biochim Biophys Acta. 1998;1442:1–19. - PubMed
-
- Coenen C, Lomax TL. Auxin-cytokinin interactions in higher plants: old problems and new tools. Trends Plant Sci. 1997;9:351–356. - PubMed
-
- Curtis IS, Power JB, Blackhall NW, de Laat AMM, Davey MR. Genotype-independent transformation of lettuce using Agrobacterium tumefaciens. J Exp Bot. 1994;279:1441–1449.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
