

The expression of alternative oxidase and uncoupling protein during fruit ripening in mango
- PMID: 11500560
- PMCID: PMC117161
- DOI: 10.1104/pp.126.4.1619
The expression of alternative oxidase and uncoupling protein during fruit ripening in mango
Abstract
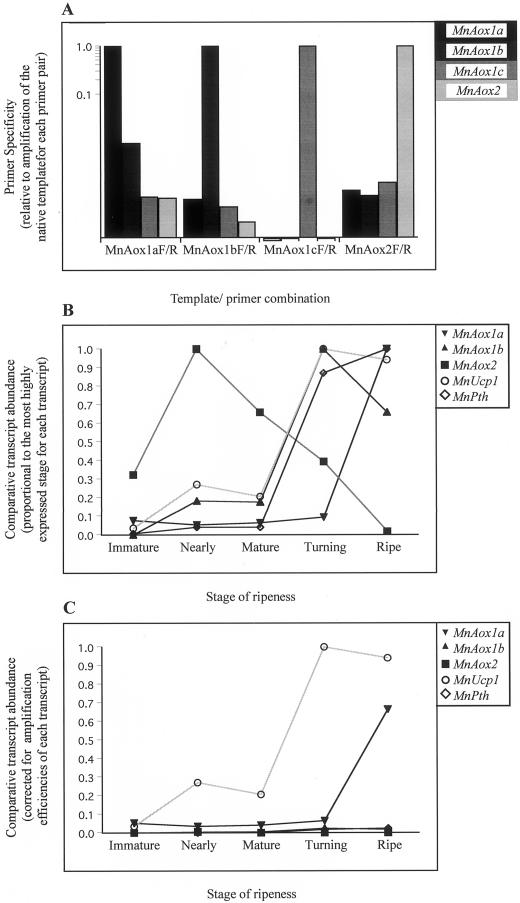
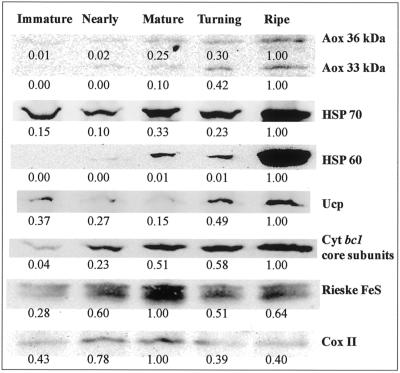
The expression of alternative oxidase (Aox) and uncoupling proteins (Ucp) was investigated during ripening in mango (Mangifera indica) and compared with the expression of peroxisomal thiolase, a previously described ripening marker in mango. The multigene family for the Aox in mango was expressed differentially during ripening. Abundance of Aox message and protein both peaked at the ripe stage. Expression of the single gene for the Ucp peaked at the turning stage and the protein abundance peaked at the ripe stage. Proteins of the cytochrome chain peaked at the mature stage of ripening. The pattern of protein accumulation suggested that increases in cytochrome chain components played an important role in facilitating the climacteric burst of respiration and that the Aox and Ucp may play a role in post-climacteric senescent processes. Because both message and protein for the Aox and Ucp increased in a similar pattern, it suggests that their expression is not controlled in a reciprocal manner but may be active simultaneously.
Figures

Similar articles
-
Involvement of AOX and UCP pathways in the post-harvest ripening of papaya fruits.J Plant Physiol. 2015 Sep 15;189:42-50. doi: 10.1016/j.jplph.2015.10.001. Epub 2015 Oct 9. J Plant Physiol. 2015. PMID: 26513459
-
Energy conservation and dissipation in mitochondria isolated from developing tomato fruit of ethylene-defective mutants failing normal ripening: the effect of ethephon, a chemical precursor of ethylene.J Bioenerg Biomembr. 2003 Apr;35(2):157-68. doi: 10.1023/a:1023750204310. J Bioenerg Biomembr. 2003. PMID: 12887014
-
Alternative oxidase from mango (Mangifera indica, L.) is differentially regulated during fruit ripening.Planta. 1995;197(4):569-76. doi: 10.1007/BF00191562. Planta. 1995. PMID: 8555961
-
Plant uncoupling mitochondrial protein and alternative oxidase: energy metabolism and stress.Biosci Rep. 2005 Jun-Aug;25(3-4):271-86. doi: 10.1007/s10540-005-2889-2. Biosci Rep. 2005. PMID: 16283557 Review.
-
Differential expression of uncoupling mitochondrial protein and alternative oxidase in the plant response to stress.J Bioenerg Biomembr. 2011 Feb;43(1):67-70. doi: 10.1007/s10863-011-9333-z. J Bioenerg Biomembr. 2011. PMID: 21253844 Review.
Cited by
-
Roles of mitochondrial energy dissipation systems in plant development and acclimation to stress.Ann Bot. 2015 Sep;116(4):583-600. doi: 10.1093/aob/mcv063. Epub 2015 May 18. Ann Bot. 2015. PMID: 25987710 Free PMC article. Review.
-
Genetic evidence for a regulatory pathway controlling alternative oxidase production in Neurospora crassa.Genetics. 2005 Jan;169(1):123-35. doi: 10.1534/genetics.104.034017. Epub 2004 Sep 30. Genetics. 2005. PMID: 15466423 Free PMC article.
-
Changes in mitochondrial electron partitioning in response to herbicides inhibiting branched-chain amino acid biosynthesis in soybean.Plant Physiol. 2003 Nov;133(3):1351-9. doi: 10.1104/pp.103.027805. Epub 2003 Oct 23. Plant Physiol. 2003. PMID: 14576285 Free PMC article.
-
Glyoxylic acid overcomes 1-MCP-induced blockage of fruit ripening in Pyrus communis L. var. 'D'Anjou'.Sci Rep. 2020 Apr 27;10(1):7084. doi: 10.1038/s41598-020-63642-z. Sci Rep. 2020. PMID: 32341384 Free PMC article.
-
Arabidopsis thaliana uncoupling proteins (AtUCPs): insights into gene expression during development and stress response and epigenetic regulation.J Bioenerg Biomembr. 2011 Feb;43(1):71-9. doi: 10.1007/s10863-011-9336-9. J Bioenerg Biomembr. 2011. PMID: 21301941 Review.
References
-
- Albury MS, Affourtit C, Moore AL. A highly conserved glutamate residue (Glu-270) is essential for plant alternative oxidase activity. J Biol Chem. 1998;273:30301–30305. - PubMed
-
- Berthold DA. Isolation of mutants of the Arabidopsis thaliana alternative oxidase (ubiquinol:oxygen oxidoreductase) resistant to salicylhydroxamic acid. Biochim Biophys Acta. 1998;1:73–83. - PubMed
-
- Biale JB, Young RE. Respiration and ripening in fruits: retrospect and prospect. In: Friend J, Rhodes MJC, editors. Recent Advances in the Biochemistry of Fruits and Vegetables. London: Academic Press; 1981. pp. 1–40.
-
- Bienengraeber M, Echtay KS, Klingenberg M. H+ transport by uncoupling protein (UCP-1) is dependent on a histidine pair, absent in UCP-2 and UCP-3. Biochemistry. 1998;37:3–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
