

CmC(A/T)GG DNA methylation in mature B cell lymphoma gene silencing
- PMID: 11504918
- PMCID: PMC56973
- DOI: 10.1073/pnas.181206898
CmC(A/T)GG DNA methylation in mature B cell lymphoma gene silencing
Abstract
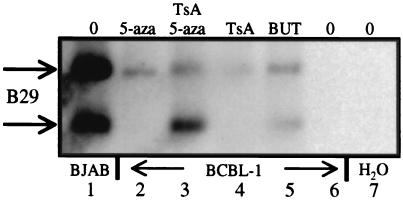
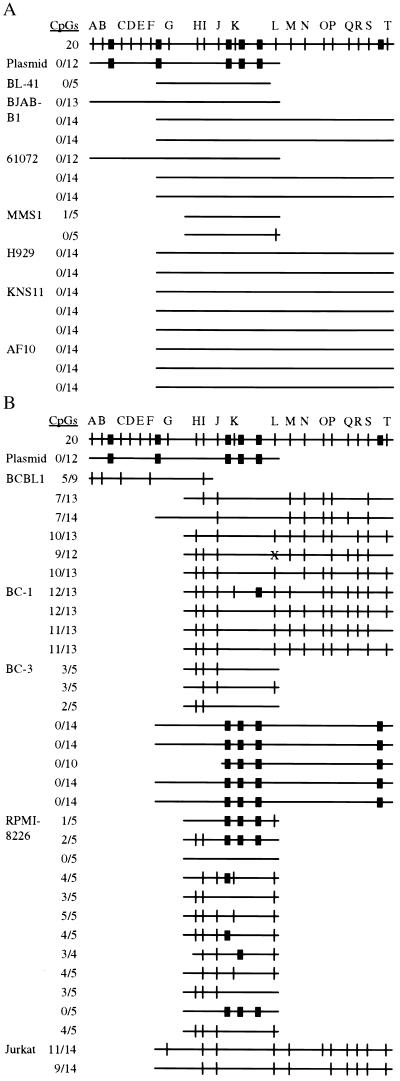
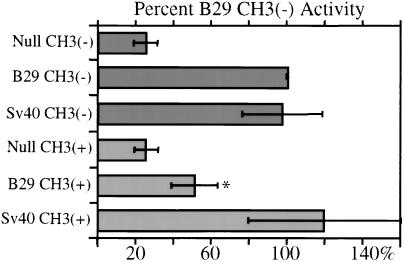
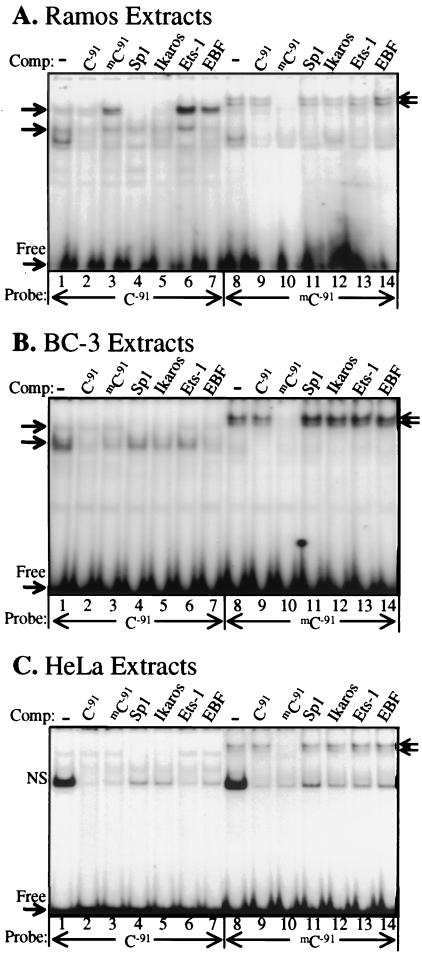
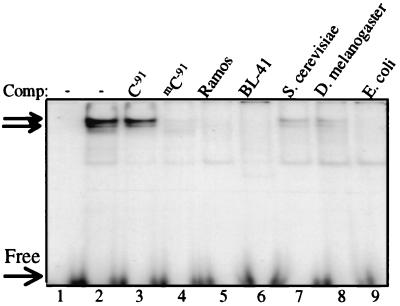
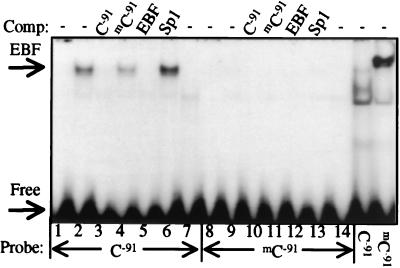
DNA methylation has been linked to gene silencing in cancer. Primary effusion lymphoma (PEL) and myeloma are lymphoid malignancies that arise from terminally differentiated B cells. Interestingly, PEL do not express immunoglobulins or most B lineage-specific genes. The B cell-specific B29 (Igbeta/CD79b) gene is silenced in PEL and some myelomas but is expressed in other normal and malignant B cells. B29 expression was reactivated in PEL by demethylating and histone deacetylase inhibiting treatments. Bisulfite sequencing revealed two types of DNA methylation in silenced B29 promoters: at conventional CpG and at CC(A/T)GG B29 promoter sites. The pattern of methylated CpG ((m)CpG) and C(m)C(A/T)GG B29 promoter methylation observed was similar to that recently reported for epigenetic silencing of an integrated retrovirus. Methylation of C(m)C(A/T)GG sites in the B29 promoter significantly repressed in vivo transcriptional activity. Also, methylation of a central conserved C(m)CTGG B29 promoter site blocked the binding of early B cell factor. This methylated motif formed DNA-protein complexes with nuclear extracts from all cell types examined. Therefore, C(m)C(A/T)GG methylation may represent an important type of epigenetic marker on mammalian DNA that impacts transcription by altering DNA-protein complex formation.
Figures
Comment in
-
C(m)C(a/t)GG methylation: a new epigenetic mark in mammalian DNA?Proc Natl Acad Sci U S A. 2001 Aug 28;98(18):10034-6. doi: 10.1073/pnas.201392598. Proc Natl Acad Sci U S A. 2001. PMID: 11526227 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
