

Failed retrograde transport of NGF in a mouse model of Down's syndrome: reversal of cholinergic neurodegenerative phenotypes following NGF infusion
- PMID: 11504920
- PMCID: PMC56979
- DOI: 10.1073/pnas.181219298
Failed retrograde transport of NGF in a mouse model of Down's syndrome: reversal of cholinergic neurodegenerative phenotypes following NGF infusion
Abstract
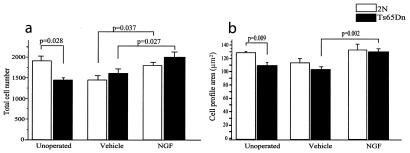
Age-related degeneration of basal forebrain cholinergic neurons (BFCNs) contributes to cognitive decline in Alzheimer's disease and Down's syndrome. With aging, the partial trisomy 16 (Ts65Dn) mouse model of Down's syndrome exhibited reductions in BFCN size and number and regressive changes in the hippocampal terminal fields of these neurons with respect to diploid controls. The changes were associated with significantly impaired retrograde transport of nerve growth factor (NGF) from the hippocampus to the basal forebrain. Intracerebroventricular NGF infusion reversed well established abnormalities in BFCN size and number and restored the deficit in cholinergic innervation. The findings are evidence that even BFCNs chronically deprived of endogenous NGF respond to an intervention that compensates for defective retrograde transport. We suggest that age-related cholinergic neurodegeneration may be a treatable disorder of failed retrograde NGF signaling.
Figures
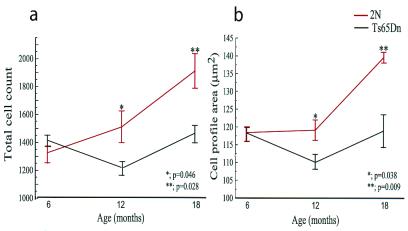
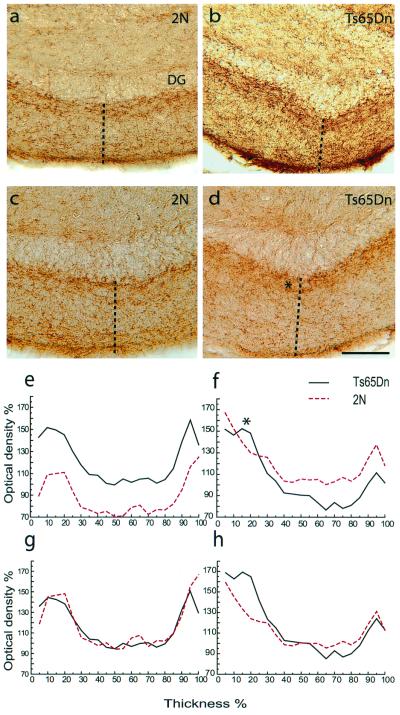
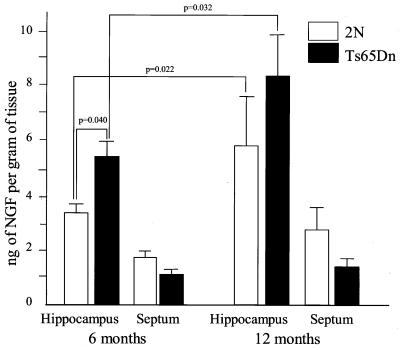
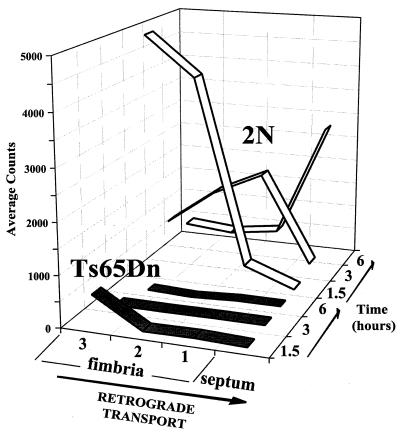
References
-
- Whitehouse P J, Price D L, Struble R G, Clark A W, Coyle J T, Delong M R. Science. 1982;215:1237–1239.
-
- Mufson E J, Bothwell M, Kordower J H. Exp Neurol. 1989;105:221–232. - PubMed
-
- Casanova M F, Walker L C, Whitehouse P J, Price D L. Ann Neurol. 1985;18:310–313. - PubMed
-
- Mann D M, Yates P O, Marcyniuk B. Neuropathol Appl Neurobiol. 1984;10:185–207. - PubMed
-
- Reeves R H, Irving N G, Moran T H, Wohn A, Kitt C, Sisodia S S, Schmidt C, Bronson R T, Davisson M T. Nat Genet. 1995;11:177–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
