

Cytochrome p450 epoxygenase metabolism of arachidonic acid inhibits apoptosis
- PMID: 11509673
- PMCID: PMC87364
- DOI: 10.1128/MCB.21.18.6322-6331.2001
Cytochrome p450 epoxygenase metabolism of arachidonic acid inhibits apoptosis
Abstract
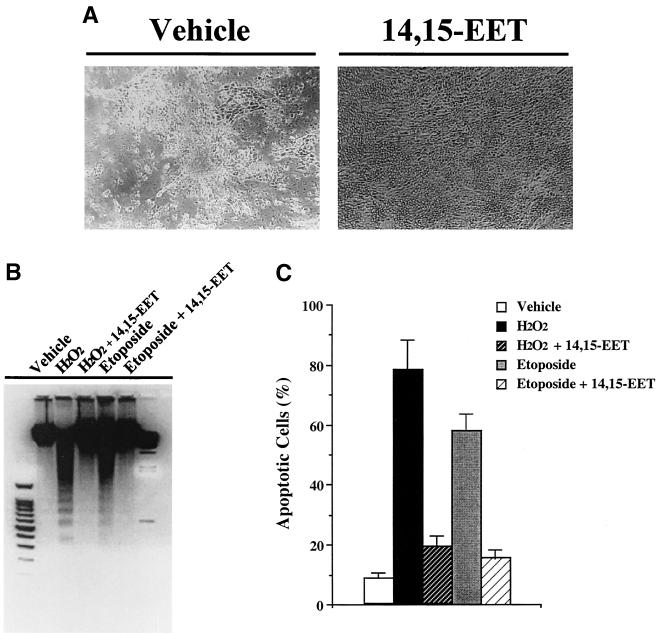
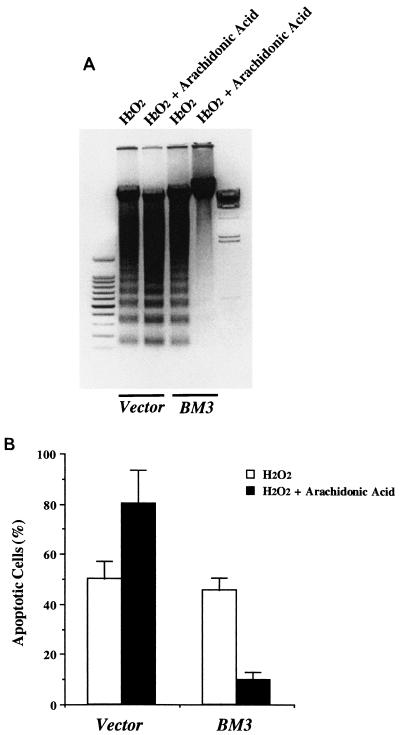
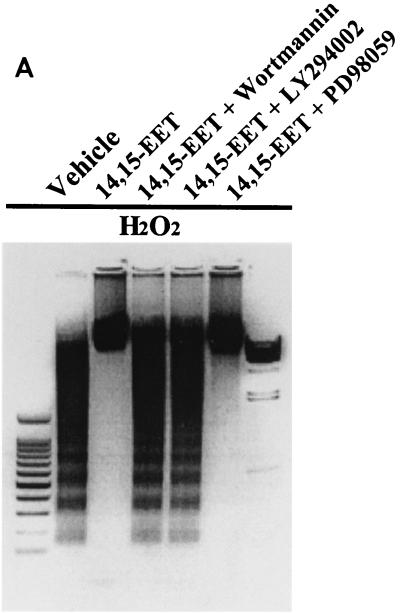
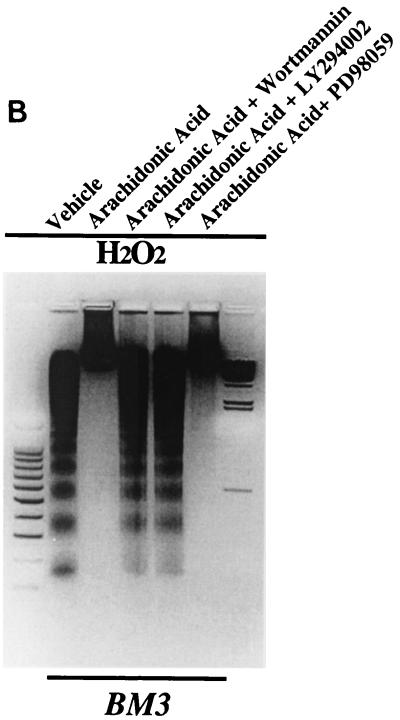
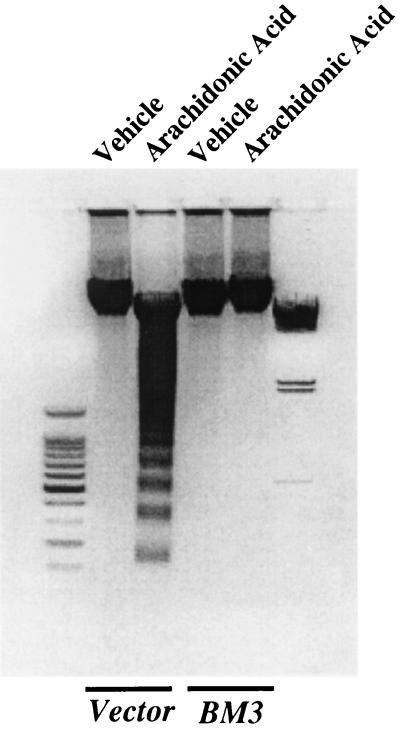
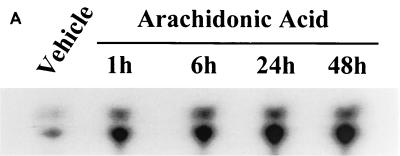
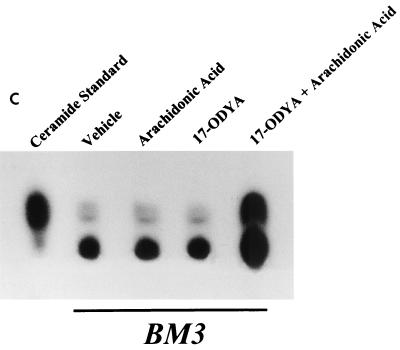
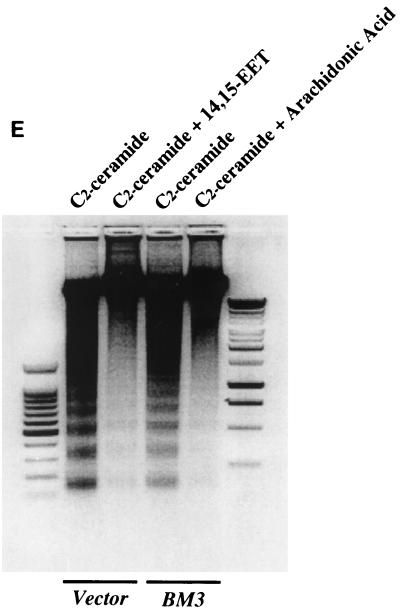
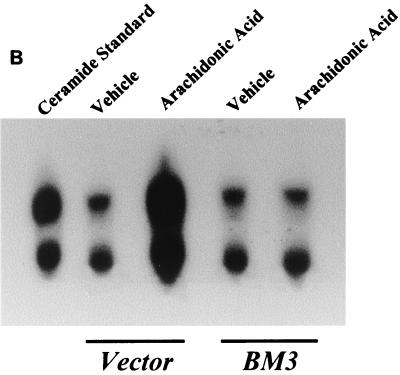
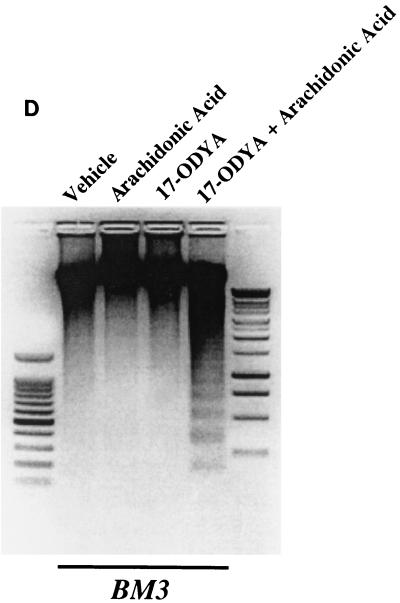
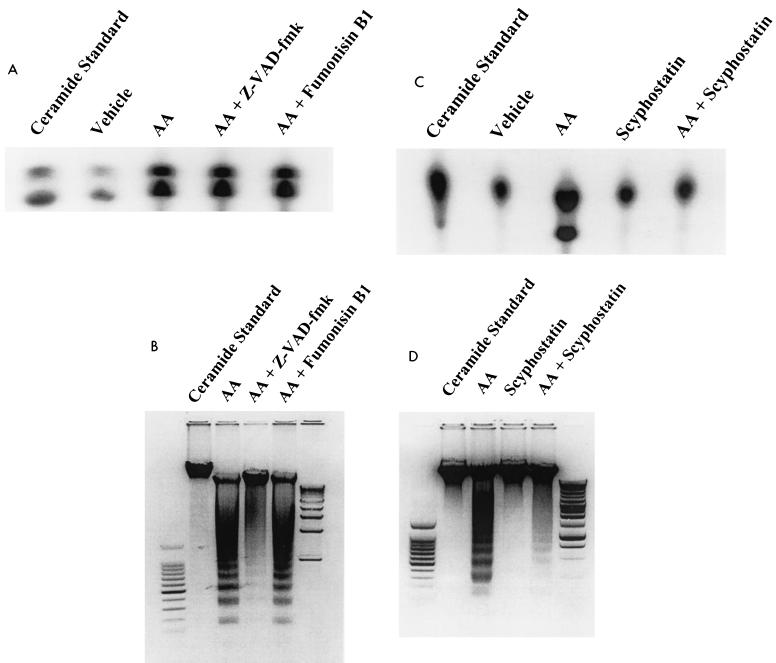
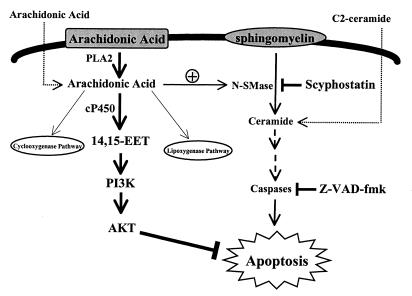
The ubiquitous cytochrome P450 hemoproteins play important functional roles in the metabolism and detoxification of foreign chemicals. However, other than established roles in cholesterol catabolism and steroid hormone biosynthesis, their cellular and/or organ physiological functions remain to be fully characterized. Here we show that the cytochrome P450 epoxygenase arachidonic acid metabolite 14,15-epoxyeicosatrienoic acid (14,15-EET) inhibits apoptosis induced by serum withdrawal, H(2)O(2), etoposide, or excess free arachidonic acid (AA), as determined by DNA laddering, Hoechst staining, and fluorescein isothiocyanate-labeled annexin V binding. In the stable transfectants (BM3 cells) expressing a mutant bacterial P450 AA epoxygenase, F87V BM3, which was genetically engineered to metabolize arachidonic acid only to 14,15-EET, AA did not induce apoptosis and protected against agonist-induced apoptosis. Ceramide assays demonstrated increased AA-induced ceramide production within 1 h and elevated ceramide levels for up to 48 h, the longest time tested, in empty-vector-transfected cells (Vector cells) but not in BM3 cells. Inhibition of cytochrome P450 activity by 17-octadecynoic acid restored AA-induced ceramide production in BM3 cells. Exogenous C2-ceramide markedly increased apoptosis in quiescent Vector cells as well as BM3 cells, and apoptosis was prevented by pretreatment of Vector cells with exogenous 14,15-EET and by pretreatment of BM3 cells with AA. The ceramide synthase inhibitor fumonisin B1 did not affect AA-induced ceramide production and apoptosis; in contrast, these effects of AA were blocked by the neutral sphingomyelinase inhibitor scyphostatin. The pan-caspase inhibitor Z-VAD-fmk had no effect on AA-induced ceramide generation but abolished AA-induced apoptosis. The antiapoptotic effects of 14,15-EET were blocked by two mechanistically and structurally distinct phosphatidylinositol-3 (PI-3) kinase inhibitors, wortmannin and LY294002, but not by the specific mitogen-activated protein kinase kinase inhibitor PD98059. Immunoprecipitation followed by an in vitro kinase assay revealed activation of Akt kinase within 10 min after 14,15-EET addition, which was completely abolished by either wortmannin or LY294002 pretreatment. In summary, the present studies demonstrated that 14,15-EET inhibits apoptosis by activation of a PI-3 kinase-Akt signaling pathway. Furthermore, cytochrome P450 epoxygenase promotes cell survival both by production of 14,15-EET and by metabolism of unesterified AA, thereby preventing activation of the neutral sphingomyelinase pathway and proapoptotic ceramide formation.
Figures

References
-
- Amsler K, Cook J S. Development of Na+-dependent hexose transport in a cultured line of porcine kidney cells. Am J Physiol. 1982;242:C94–C101. - PubMed
-
- Ara G, Teicher B A. Cyclooxygenase and lipoxygenase inhibitors in cancer therapy. Prostaglandins Leukot Essent Fatty Acids. 1996;54:3–16. - PubMed
-
- Bonvalet J P, Pradelles P, Farman N. Segmental synthesis and actions of prostaglandins along the nephron. Am J Physiol. 1987;253:F377–F387. - PubMed
-
- Bortuzzo C, Hanif R, Kashfi K, Staiano-Coico L, Shiff S J, Rigas B. The effect of leukotrienes B and selected HETEs on the proliferation of colon cancer cells. Biochim Biophys Acta. 1996;1300:240–246. - PubMed
-
- Burgering B M, Coffer P J. Protein kinase B (c-Akt) in phosphatidylinositol-3-OH kinase signal transduction. Nature. 1995;376:599–602. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous