

A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae
- PMID: 11511543
- PMCID: PMC312755
- DOI: 10.1101/gad.204401
A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae
Abstract
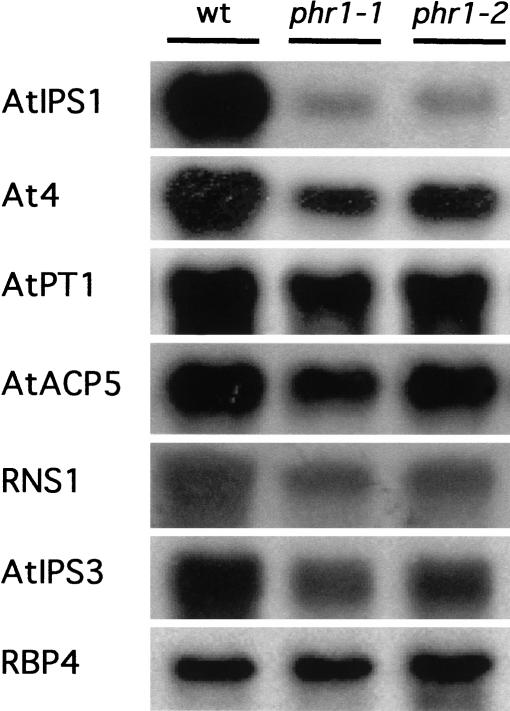
Plants have evolved a number of adaptive responses to cope with growth in conditions of limited phosphate (Pi) supply involving biochemical, metabolic, and developmental changes. We prepared an EMS-mutagenized M(2) population of an Arabidopsis thaliana transgenic line harboring a reporter gene specifically responsive to Pi starvation (AtIPS1::GUS), and screened for mutants altered in Pi starvation regulation. One of the mutants, phr1 (phosphate starvation response 1), displayed reduced response of AtIPS1::GUS to Pi starvation, and also had a broad range of Pi starvation responses impaired, including the responsiveness of various other Pi starvation-induced genes and metabolic responses, such as the increase in anthocyanin accumulation. PHR1 was positionally cloned and shown be related to the PHOSPHORUS STARVATION RESPONSE 1 (PSR1) gene from Chlamydomonas reinhardtii. A GFP::PHR1 protein fusion was localized in the nucleus independently of Pi status, as is the case for PSR1. PHR1 is expressed in Pi sufficient conditions and, in contrast to PSR1, is only weakly responsive to Pi starvation. PHR1, PSR1, and other members of the protein family share a MYB domain and a predicted coiled-coil (CC) domain, defining a subtype within the MYB superfamily, the MYB-CC family. Therefore, PHR1 was found to bind as a dimer to an imperfect palindromic sequence. PHR1-binding sequences are present in the promoter of Pi starvation-responsive structural genes, indicating that this protein acts downstream in the Pi starvation signaling pathway.
Figures







References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Ames BN. Assay of inorganic phosphate, total phosphate and phosphatases. Methods Enzymol. 1966;8:115–118.
-
- Bariola PA, Howard CJ, Taylor CB, Verburg MT, Jaglan VD, Green PJ. The Arabidopsis ribonuclease gene RSN1 is tightly controlled in response to phosphate limitation. Plant J. 1994;6:673–685. - PubMed
-
- Bates TR, Lynch JP. Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability. Plant Cell Environ. 1996;19:529–538.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous