

Mutational analysis of the Rhizobium lupini H13-3 and Sinorhizobium meliloti flagellin genes: importance of flagellin A for flagellar filament structure and transcriptional regulation
- PMID: 11514517
- PMCID: PMC95416
- DOI: 10.1128/JB.183.18.5334-5342.2001
Mutational analysis of the Rhizobium lupini H13-3 and Sinorhizobium meliloti flagellin genes: importance of flagellin A for flagellar filament structure and transcriptional regulation
Abstract
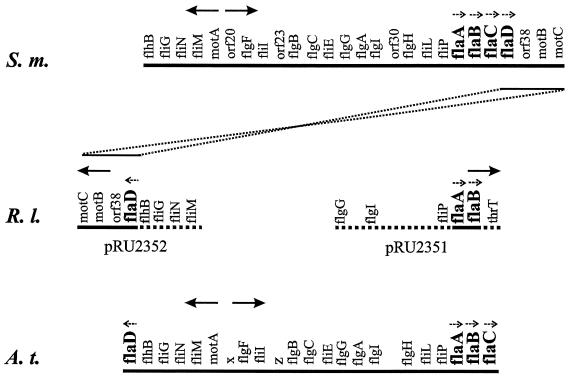
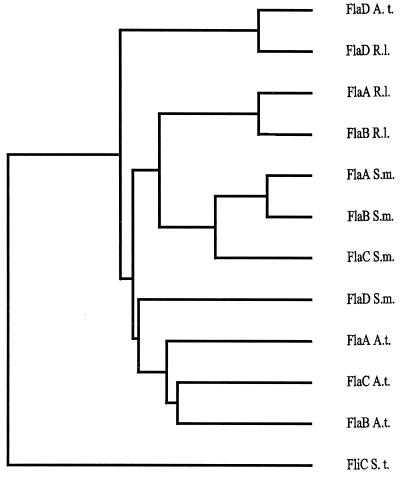
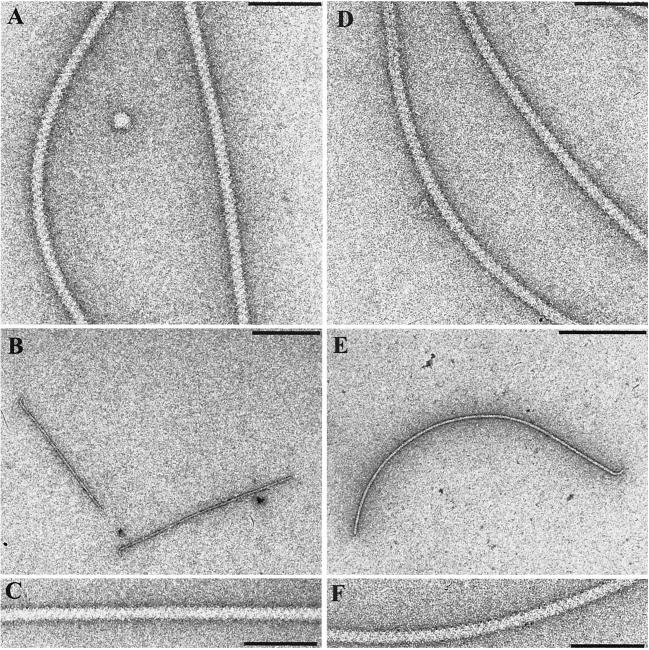
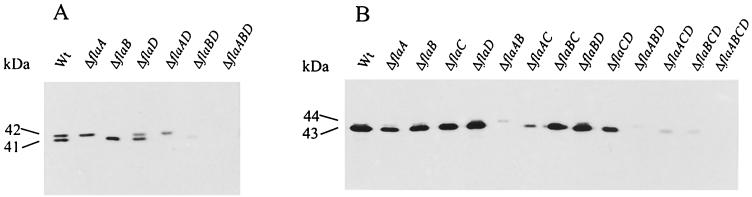
Complex flagellar filaments are unusual in their fine structure composed of flagellin dimers, in their right-handed helicity, and in their rigidity, which prevents a switch of handedness. The complex filaments of Rhizobium lupini H13-3 and those of Sinorhizobium meliloti are composed of three and four flagellin (Fla) subunits, respectively. The Fla-encoding genes, named flaA through flaD, are separately transcribed from sigma(28)-specific promoters. Mutational analysis of the fla genes revealed that, in both species, FlaA is the principal flagellin and that FlaB, FlaC, and FlaD are secondary. FlaA and at least one secondary Fla protein are required for assembling a functional flagellar filament. Western analysis revealed a ratio close to 1 of FlaA to the secondary Fla proteins (= FlaX) present in wild-type extracts, suggesting that the complex filament is assembled from FlaA-FlaX heterodimers. Whenever a given mutant combination of Fla prevented the assemblage of an intact filament, the biosynthesis of flagellin decreased dramatically. As shown in S. meliloti by reporter gene analysis, it is the transcription of flaA, but not of flaB, flaC, or flaD, that was down-regulated by such abortive combinations of Fla proteins. This autoregulation of flaA is unusual. We propose that any combination of Fla subunits incapable of assembling an intact filament jams the flagellar export channel and thus prevents the escape of an (as yet unidentified) anti-sigma(28) factor that antagonizes the sigma(28)-dependent transcription of flaA.
Figures


References
-
- Aizawa S-I. Encyclopedia of microbiology. 2nd ed. Vol. 2. New York, N.Y: Academic Press, Inc.; 2000. Flagella; pp. 380–389.
-
- Chesnokova O, Coutinho J B, Khan I H, Mikhail M S, Kado C I. Characterization of flagella genes of Agrobacterium tumefaciens, and the effect of a bald strain on virulence. Mol Microbiol. 1997;23:579–590. - PubMed
-
- Cohen-Krausz S, Trachtenberg S. Helical perturbations of the flagellar filament: Rhizobium lupini H13–3 at 13 Å resolution. J Struct Biol. 1998;122:267–282. - PubMed
-
- Deakin W J, Parker V E, Wright E L, Ashcroft K J, Loake G J, Shaw C H. Agrobacterium tumefaciens possesses a fourth flagellin gene located in a large gene cluster concerned with flagellar structure, assembly and motility. Microbiology. 1999;145:1397–1407. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
