

Hop resistance in the beer spoilage bacterium Lactobacillus brevis is mediated by the ATP-binding cassette multidrug transporter HorA
- PMID: 11514522
- PMCID: PMC95421
- DOI: 10.1128/JB.183.18.5371-5375.2001
Hop resistance in the beer spoilage bacterium Lactobacillus brevis is mediated by the ATP-binding cassette multidrug transporter HorA
Abstract
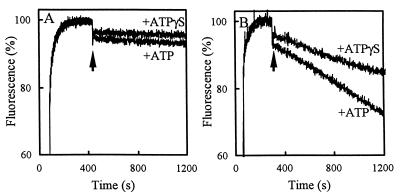
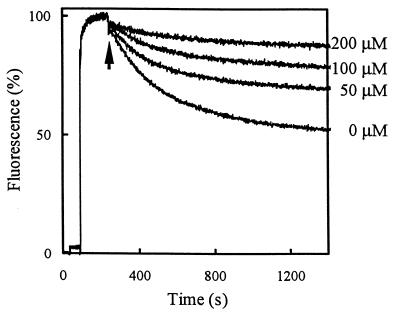
Lactobacillus brevis is a major contaminant of spoiled beer. The organism can grow in beer in spite of the presence of antibacterial hop compounds that give the beer a bitter taste. The hop resistance in L. brevis is, at least in part, dependent on the expression of the horA gene. The deduced amino acid sequence of HorA is 53% identical to that of LmrA, an ATP-binding cassette multidrug transporter in Lactococcus lactis. To study the role of HorA in hop resistance, HorA was functionally expressed in L. lactis as a hexa-histidine-tagged protein using the nisin-controlled gene expression system. HorA expression increased the resistance of L. lactis to hop compounds and cytotoxic drugs. Drug transport studies with L. lactis cells and membrane vesicles and with proteoliposomes containing purified HorA protein identified HorA as a new member of the ABC family of multidrug transporters.
Figures
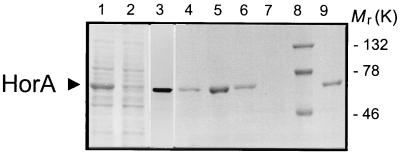
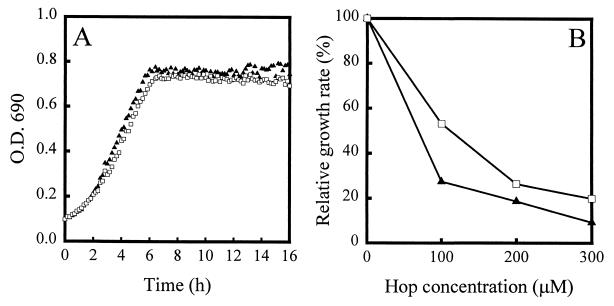
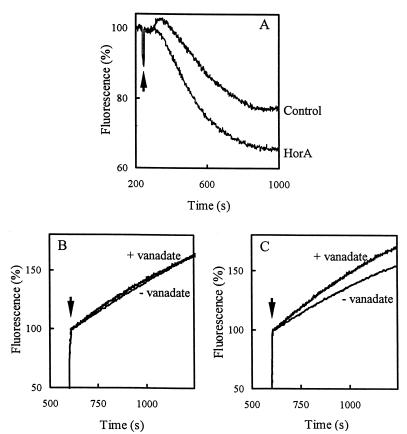
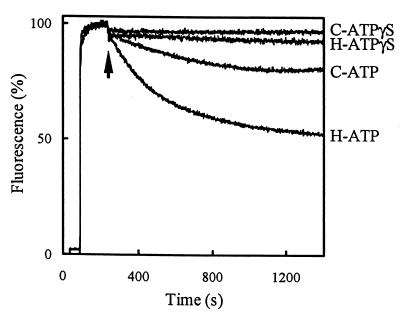
References
-
- Davies J. Inactivation of antibiotics and the dissemination of resistance genes. Science. 1994;264:375–382. - PubMed
-
- Higgins C F. ABC transporters: from microorganisms to man. Annu Rev Cell Biol. 1992;8:67–113. - PubMed
-
- Margolles M, Putman M, van Veen H W, Konings W N. The purified and functionally reconstituted multidrug transporter LmrA of Lactococcus lactis mediates the transbilayer movement of specific fluorescent phospholipids. Biochemistry. 1999;38:16298–16306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
