

ER beta inhibits proliferation and invasion of breast cancer cells
- PMID: 11517191
- PMCID: PMC2040491
- DOI: 10.1210/endo.142.9.8395
ER beta inhibits proliferation and invasion of breast cancer cells
Abstract
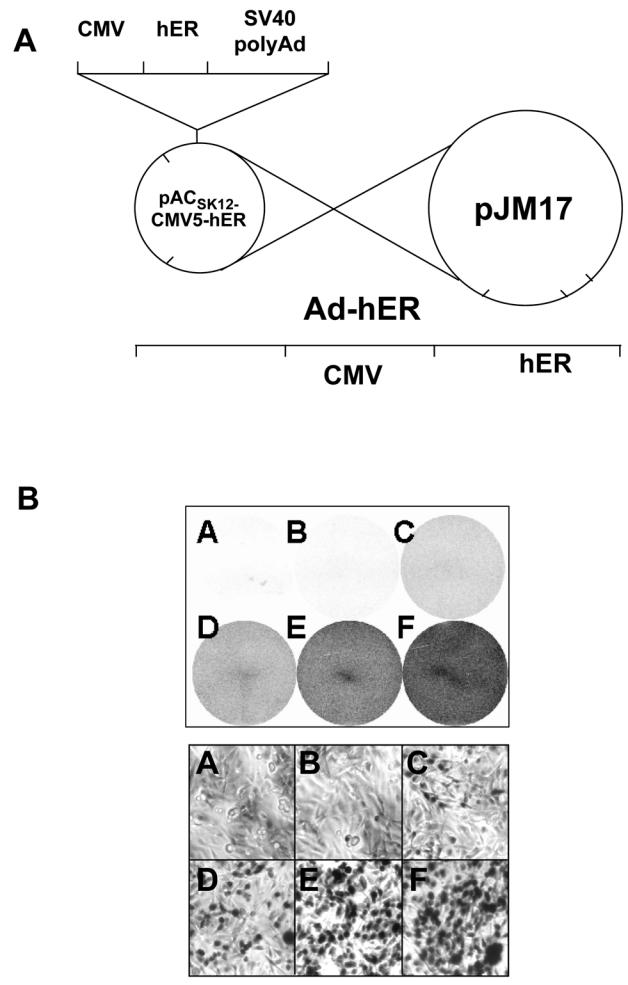
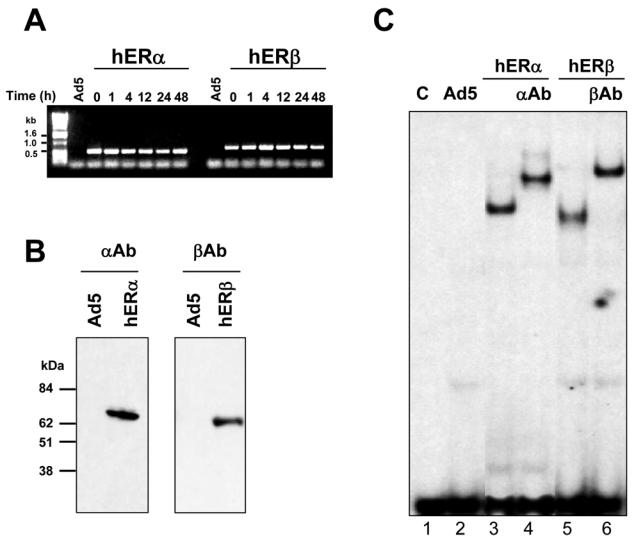
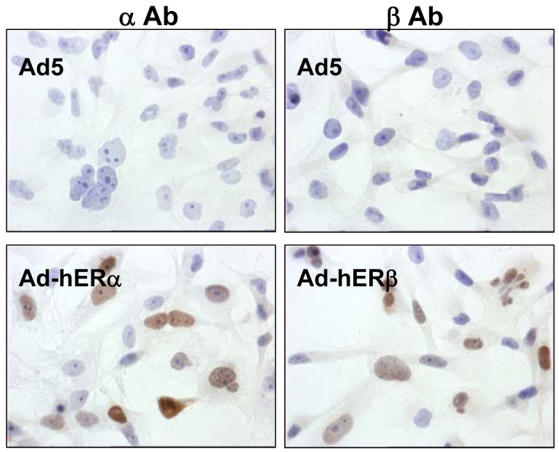
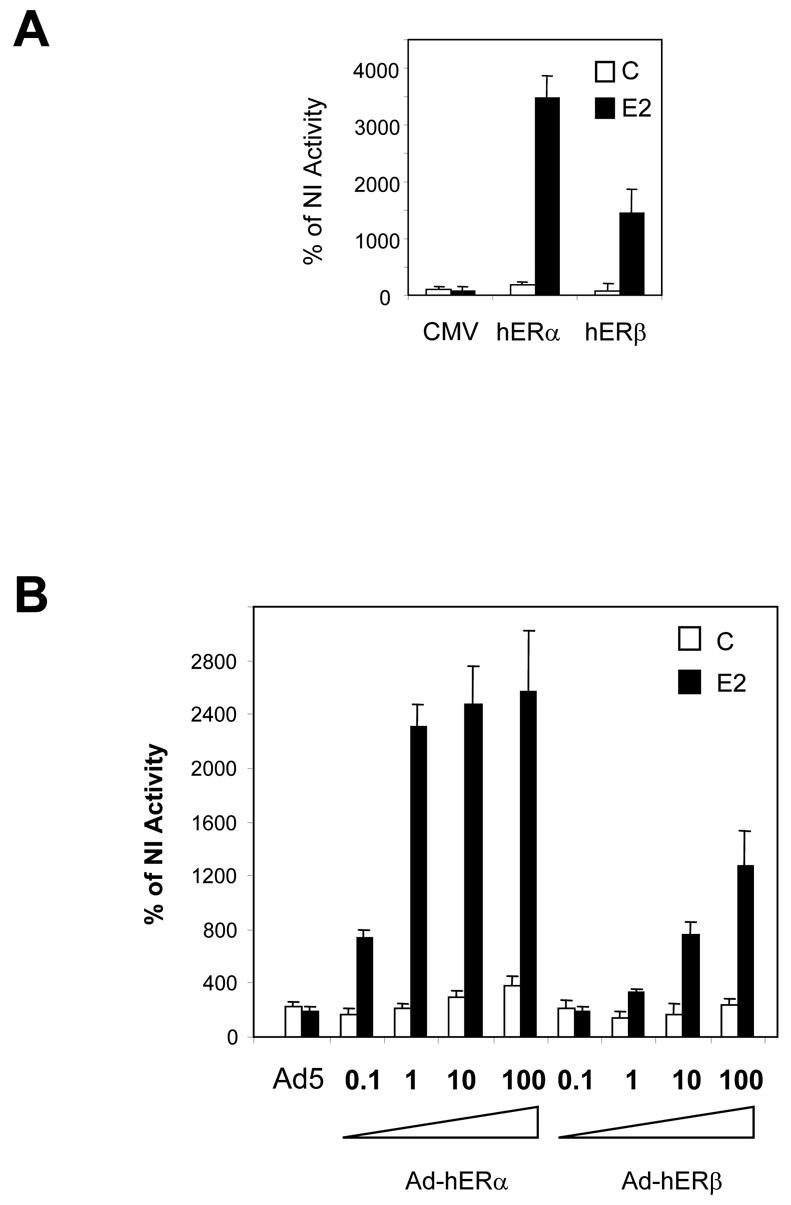
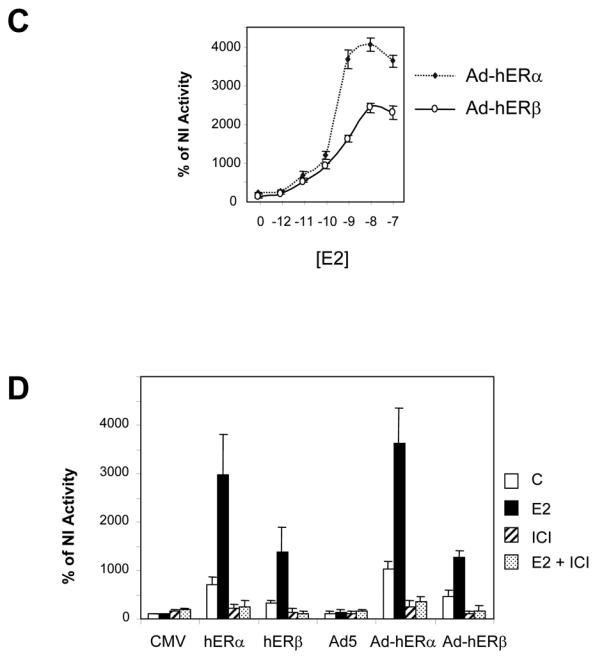
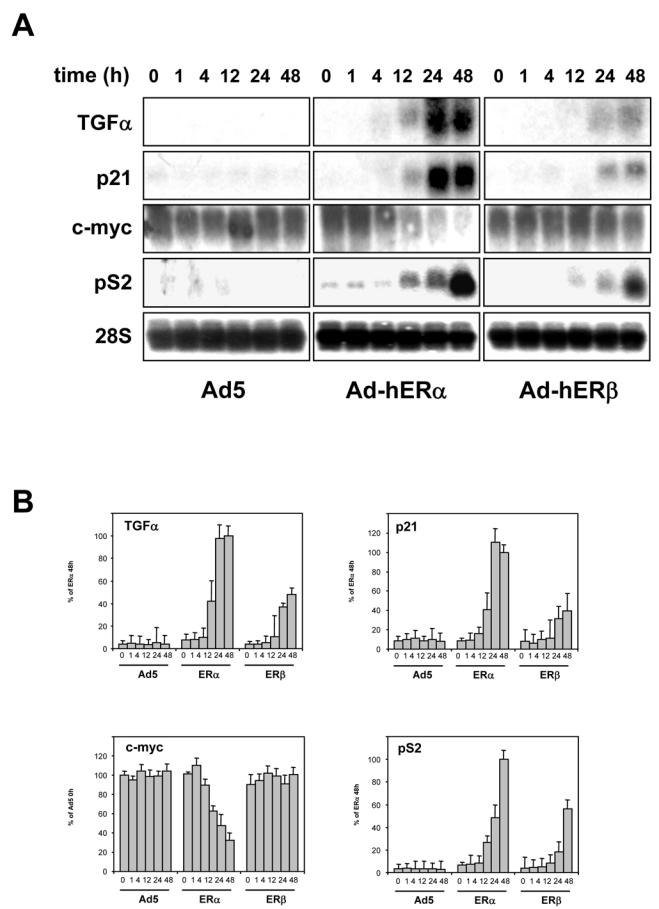
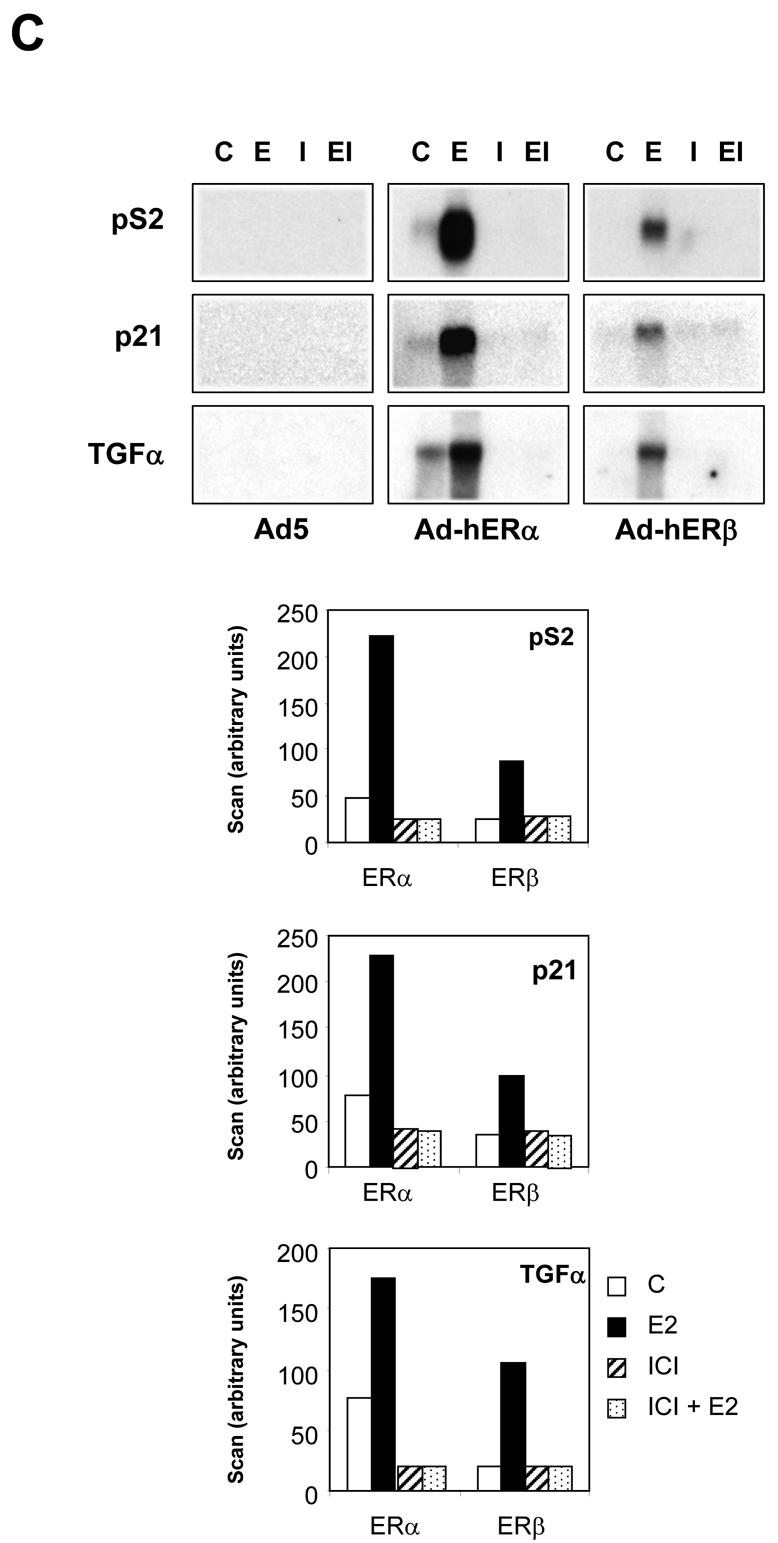
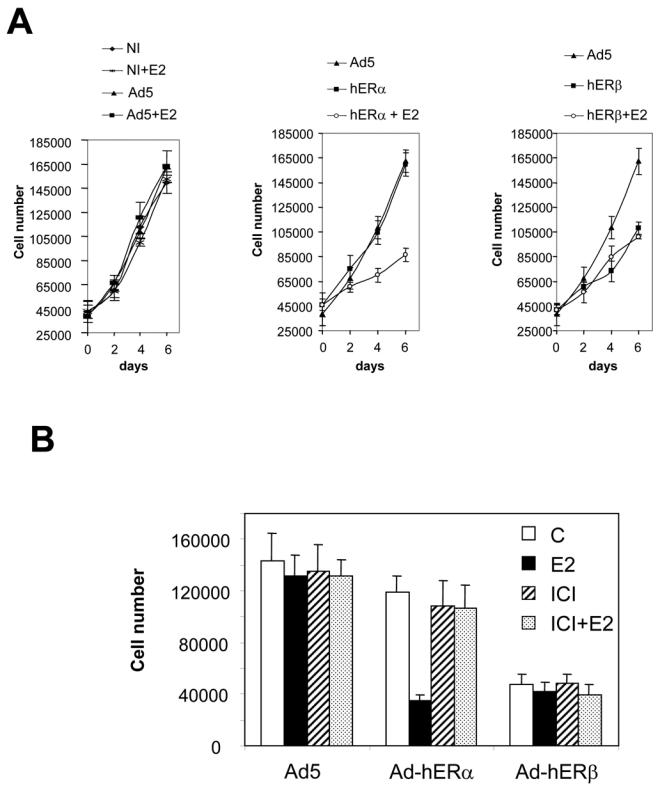
Recent studies indicate that the expression of ER beta in breast cancer is lower than in the normal breast, suggesting that ER beta could play an important role in carcinogenesis. To investigate this hypothesis, we engineered ER-negative MDA-MB-231 (human breast cancer cells) to reintroduce either ER alpha or ER beta protein with an adenoviral vector. In these cells, ER beta (as ER alpha) expression was monitored using RT-PCR and Western blot. ER beta protein was localized in the nucleus (immunocytochemistry) and able to transactivate estrogen-responsive reporter constructs in the presence of E2. ER beta and ER alpha induced the expression of several endogenous genes such as pS2, TGF alpha, or the cyclin kinase inhibitor p21 but, in contrast to ER alpha, ER beta was unable to regulate c-myc proto-oncogene expression. The pure antiestrogen ICI 164, 384 completely blocked ER alpha and ER beta estrogen-induced activities. ER beta inhibited MDA-MB-231 cell proliferation in a ligand-independent manner, whereas ER alpha inhibition of proliferation is hormone dependent. Moreover, ER beta and ER alpha decreased cell motility and invasion. Our data bring the first evidence that ER beta is an important modulator of proliferation and invasion of breast cancer cells and support the hypothesis that the loss of ER beta expression could be one of the events leading to the development of breast cancer.
Figures
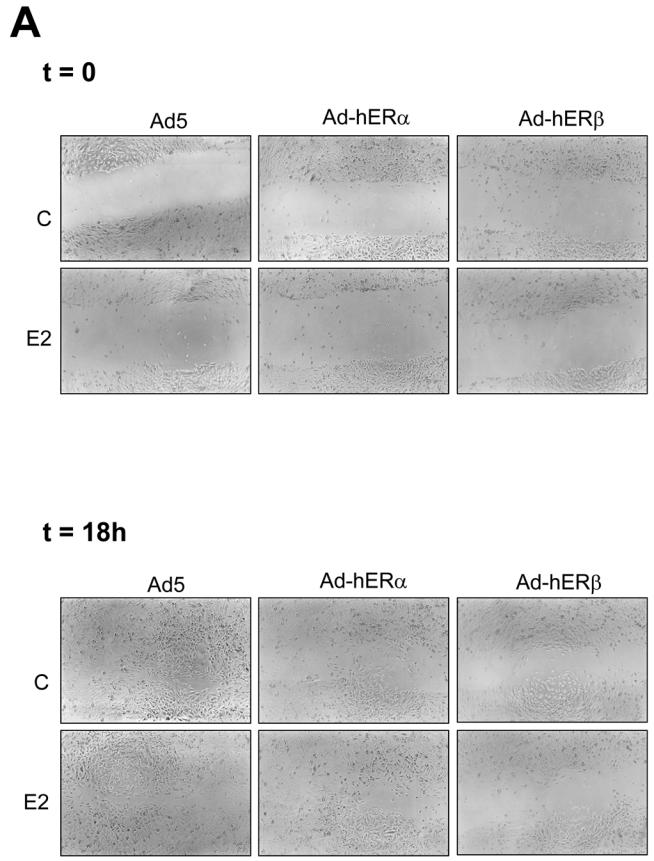
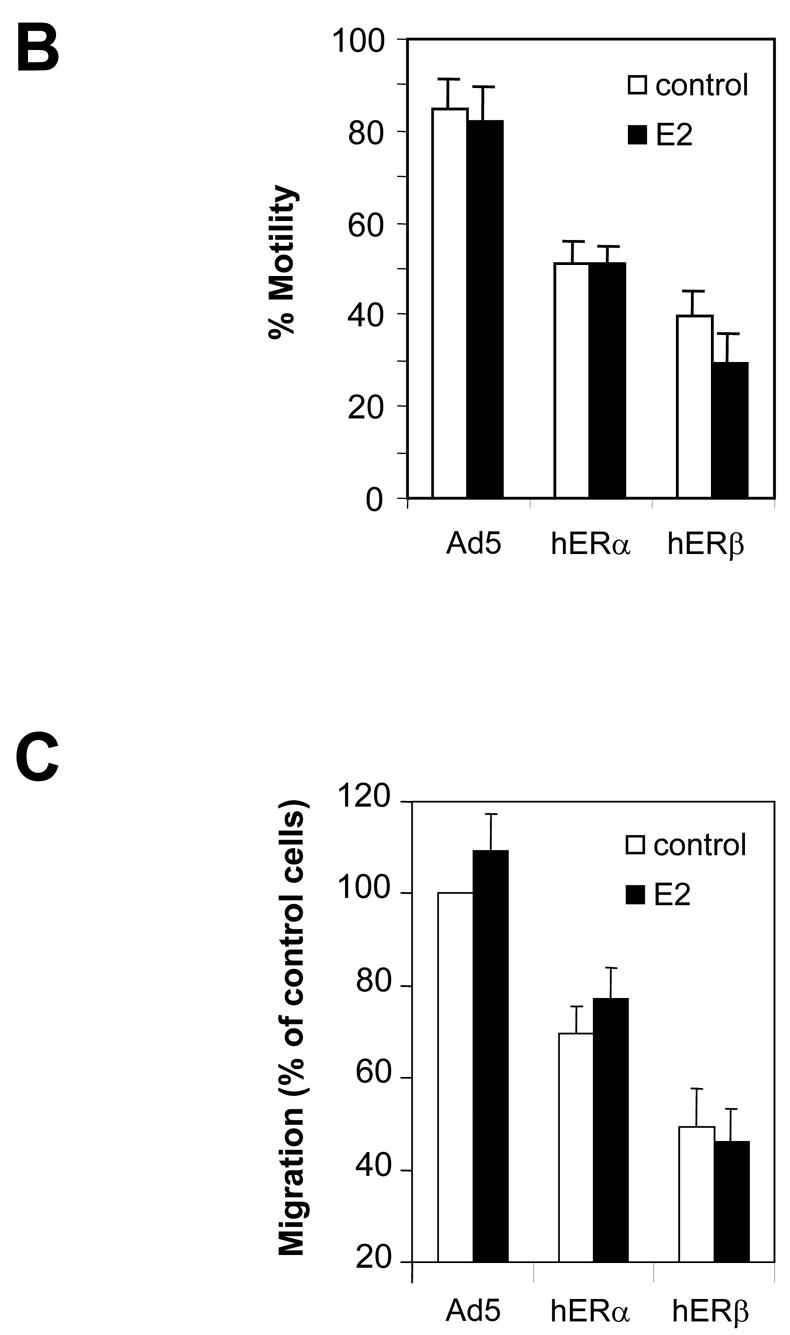
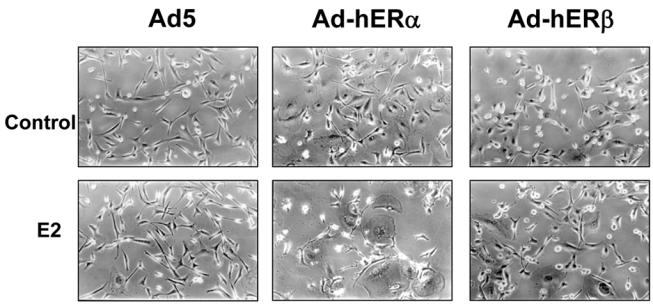
References
-
- Gustafsson JA. Estrogen receptor beta--a new dimension in estrogen mechanism of action. J Endocrinol. 1999;163:379–83. - PubMed
-
- Russo J, Russo IH. Differentiation and breast cancer. Medicina (B Aires) 1997;57:81–91. - PubMed
-
- Haslam SZ, Counterman LJ, Nummy KA. Effects of epidermal growth factor, estrogen, and progestin on DNA synthesis in mammary cells in vivo are determined by the developmental state of the gland. J Cell Physiol. 1993;155:72–8. - PubMed
-
- Hilakivi-Clarke L, Clarke R, Lippman M. The influence of maternal diet on breast cancer risk among female offspring. Nutrition. 1999;15:392–401. - PubMed
-
- Liao DZ, Pantazis CG, Hou X, Li SA. Promotion of estrogen-induced mammary gland carcinogenesis by androgen in the male Noble rat: probable mediation by steroid receptors. Carcinogenesis. 1998;19:2173–80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
