

Chronic blockade of glutamate receptors enhances presynaptic release and downregulates the interaction between synaptophysin-synaptobrevin-vesicle-associated membrane protein 2
- PMID: 11517248
- PMCID: PMC6763110
- DOI: 10.1523/JNEUROSCI.21-17-06588.2001
Chronic blockade of glutamate receptors enhances presynaptic release and downregulates the interaction between synaptophysin-synaptobrevin-vesicle-associated membrane protein 2
Abstract
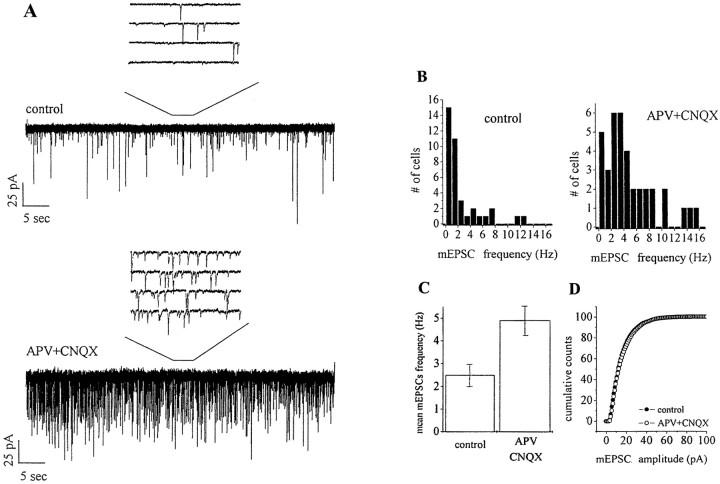
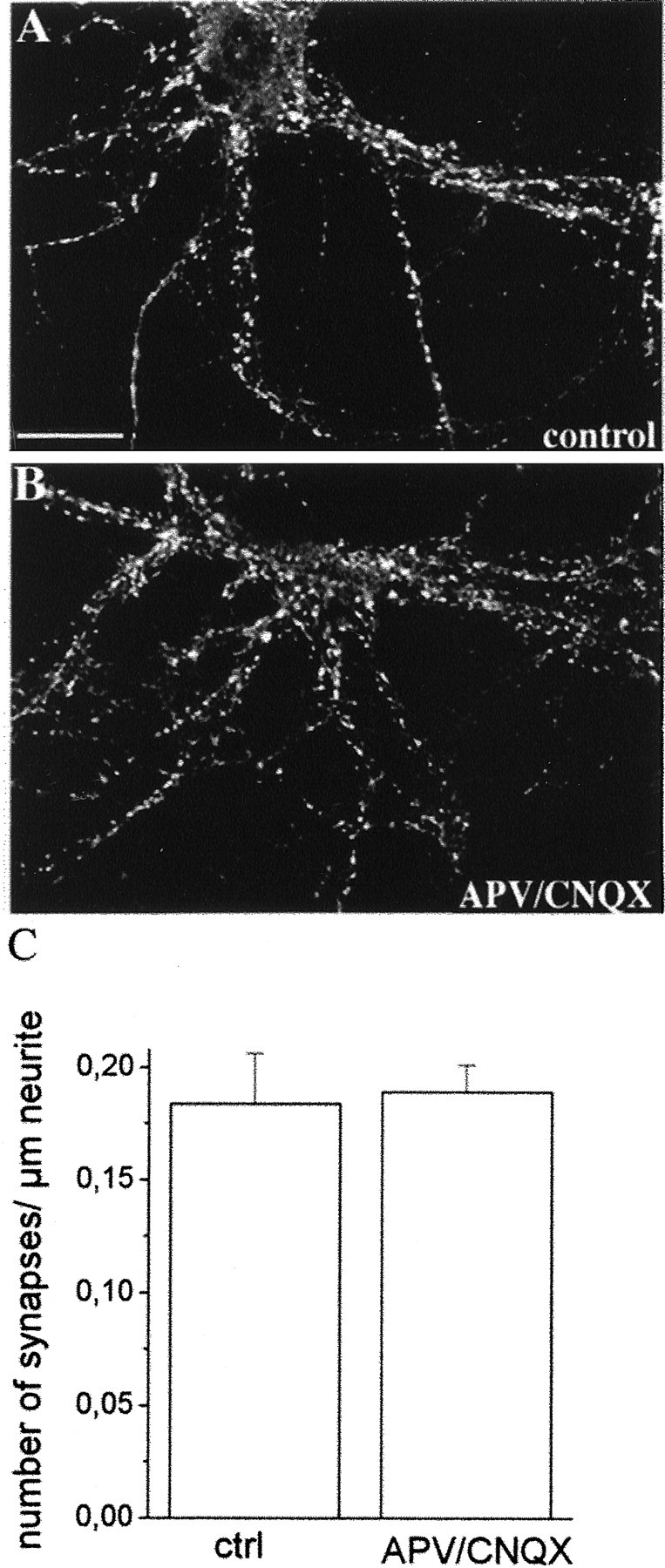
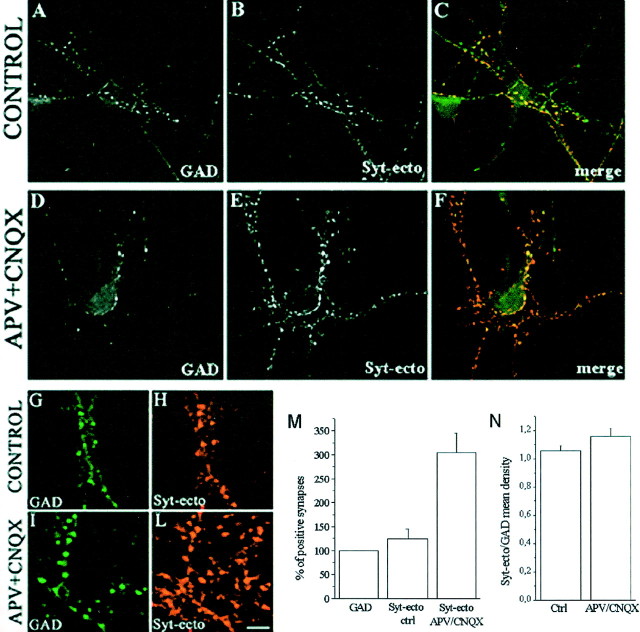
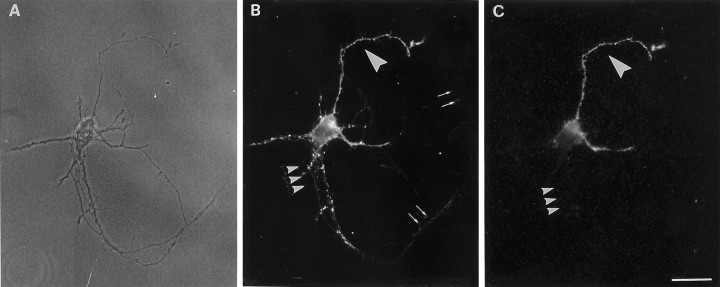
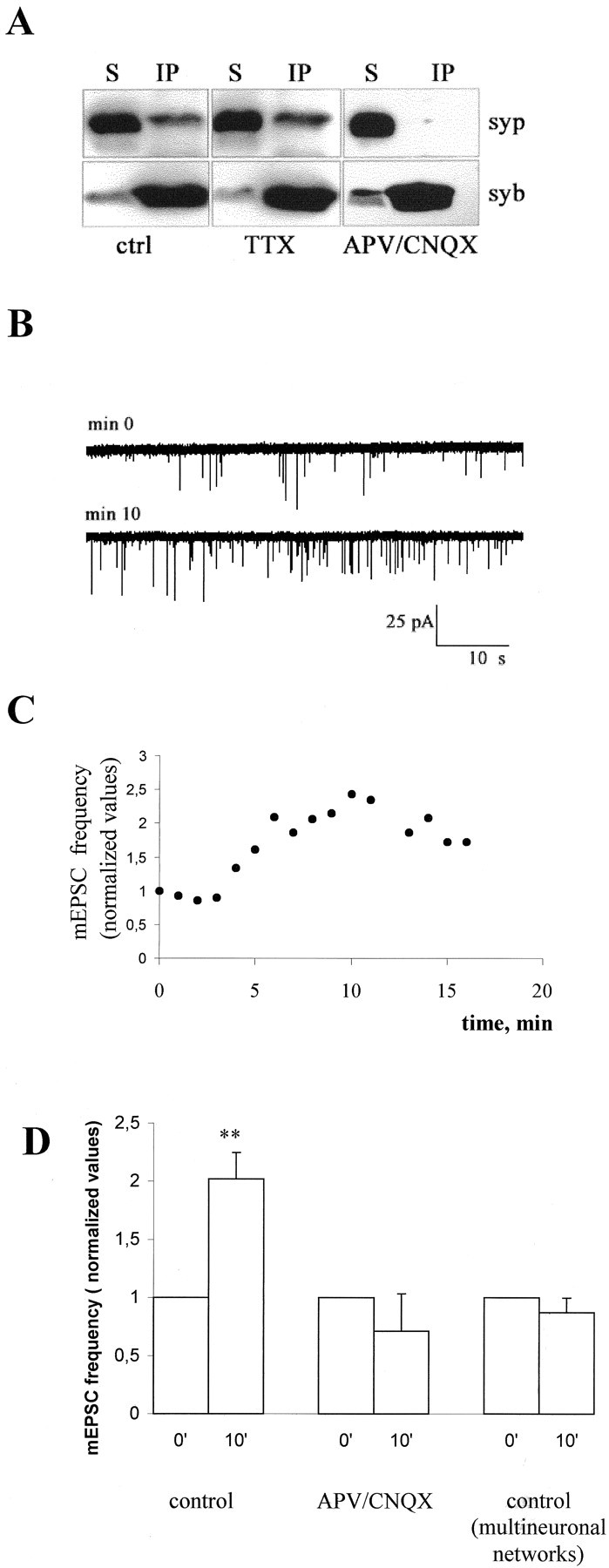
During development of neuronal circuits, presynaptic and postsynaptic functions are adjusted in concert, to optimize interneuronal signaling. We have investigated whether activation of glutamate receptors affects presynaptic function during synapse formation, when constitutive synaptic vesicle recycling is downregulated. Using primary cultures of hippocampal neurons as a model system, we have found that chronic exposure to both NMDA and non-NMDA glutamate receptor blockers during synaptogenesis produces an increase in miniature EPSC (mEPSC) frequency, with no significant changes in mEPSC amplitude or in the number of synapses. Enhanced synaptic vesicle recycling, selectively in glutamatergic nerve terminals, was confirmed by the increased uptake of antibodies directed against the lumenal domain of synaptotagmin. No increased uptake was detected in neuronal cultures grown in the chronic presence of TTX, speaking against an indirect effect caused by decreased electrical activity. Enhanced mEPSC frequency correlated with a reduction of synaptophysin-synaptobrevin-vesicle-associated membrane protein 2 (VAMP2) complexes detectable by immunoprecipitation. Intracellular perfusion with a peptide that inhibits the binding of synaptophysin to synaptobrevin-VAMP2 induced a remarkable increase of mEPSC frequency in control but not in glutamate receptor blocker-treated neurons. These findings suggest that activation of glutamate receptors plays a role in the downregulation of the basal rate of synaptic vesicle recycling that accompanies synapse formation. They also suggest that one of the mechanisms through which this downregulation is achieved is an increased interaction of synaptophysin with synaptobrevin-VAMP2.
Figures

References
-
- Ahmari SE, Buchanan J, Smith SJ. Assembly of presynaptic active zones from cytoplasmic transport packets. Nat Neurosci. 2000;3:445–451. - PubMed
-
- Becher A, Drenckhahn A, Pahner I, Ahnert-Hilger G. The synaptophysin-synaptobrevin complex is developmentally upregulated in cultivated neurons but is absent in neuroendocrine cells. Eur J Cell Biol. 1999b;78:650–656. - PubMed
-
- Benson DL, Watkins FH, Stewaed O, Banker GA. Characterization of GABAergic neurons in hippocampal cell cultures. J Neurocytol. 1994;23:279–295. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources