

Non-enzymatic glycosylation of a type I collagen matrix: effects on osteoblastic development and oxidative stress
- PMID: 11518540
- PMCID: PMC37548
- DOI: 10.1186/1471-2121-2-16
Non-enzymatic glycosylation of a type I collagen matrix: effects on osteoblastic development and oxidative stress
Abstract
Background: The tissue accumulation of protein-bound advanced glycation endproducts (AGE) may be involved in the etiology of diabetic chronic complications, including osteopenia. The aim of this study was to investigate the effect of an AGE-modified type I collagen substratum on the adhesion, spreading, proliferation and differentiation of rat osteosarcoma UMR106 and mouse non-transformed MC3T3E1 osteoblastic cells. We also studied the role of reactive oxygen species (ROS) and nitric oxide synthase (NOS) expression on these AGE-collagen mediated effects.
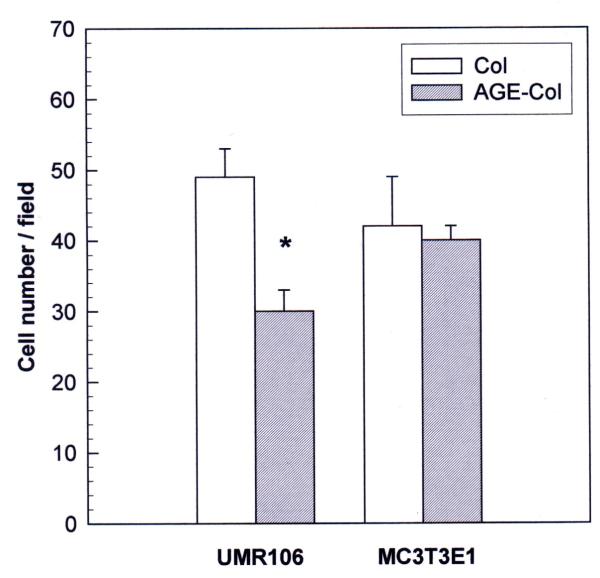
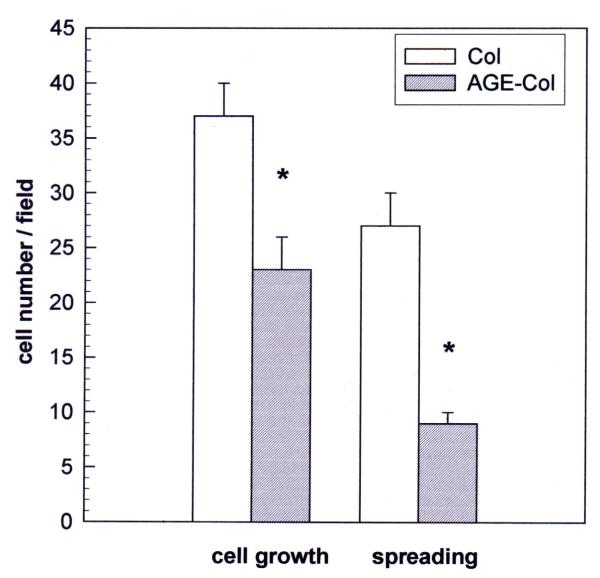
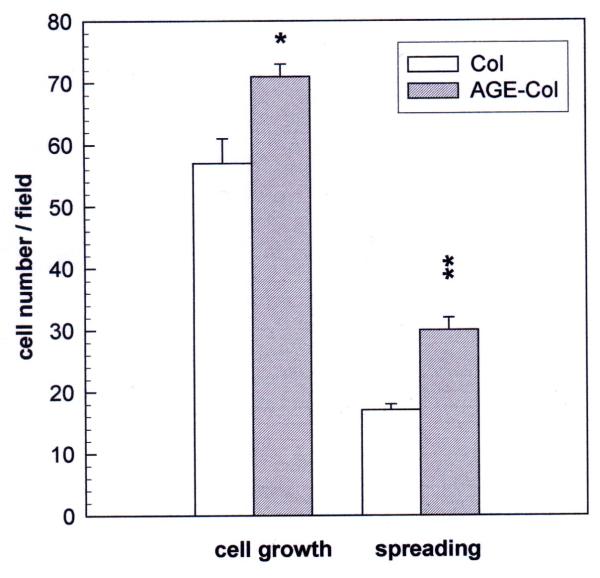
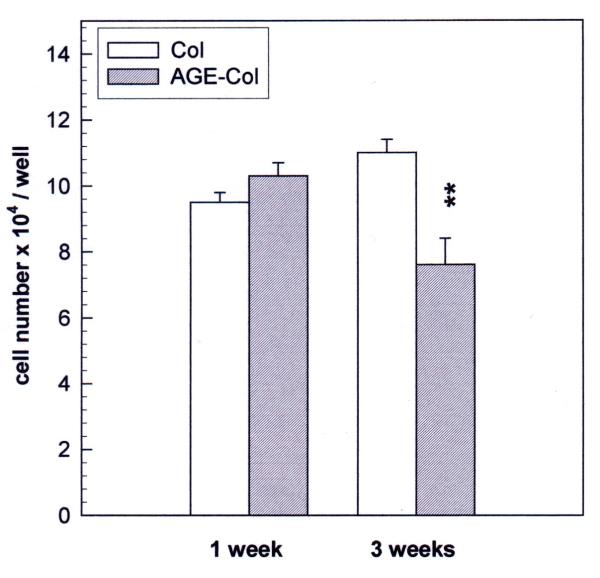
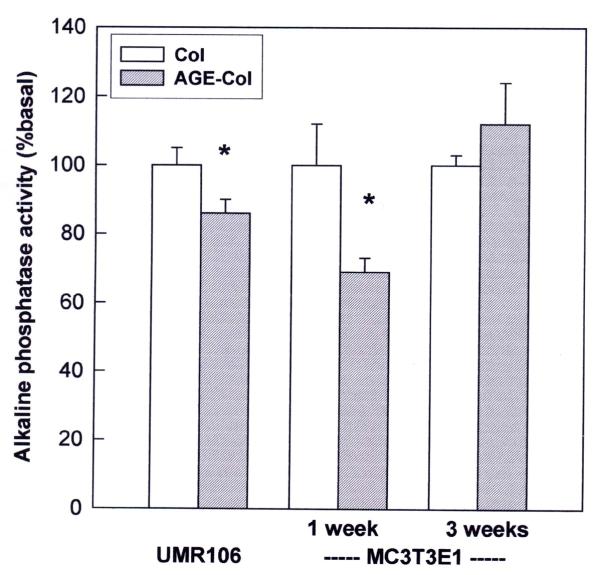
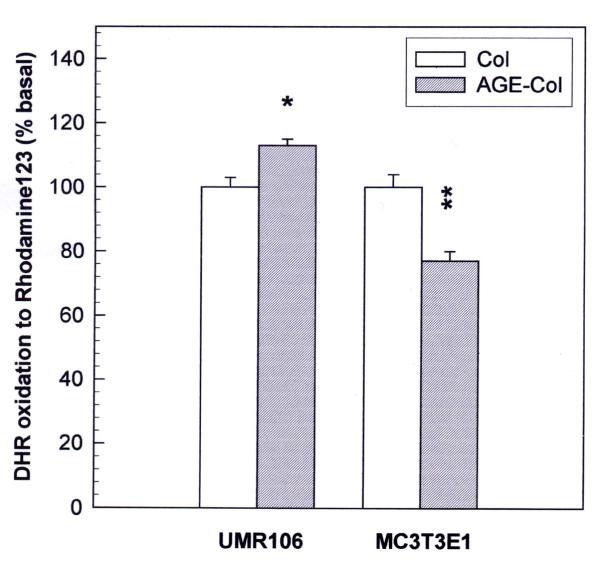
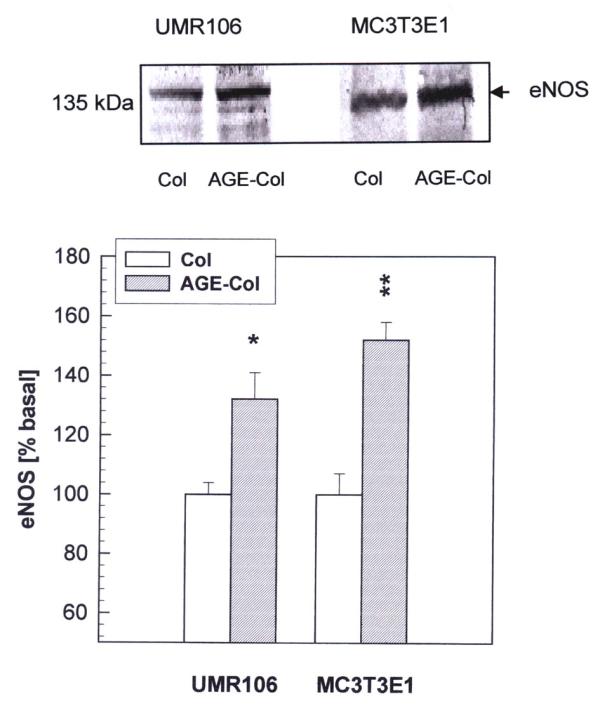
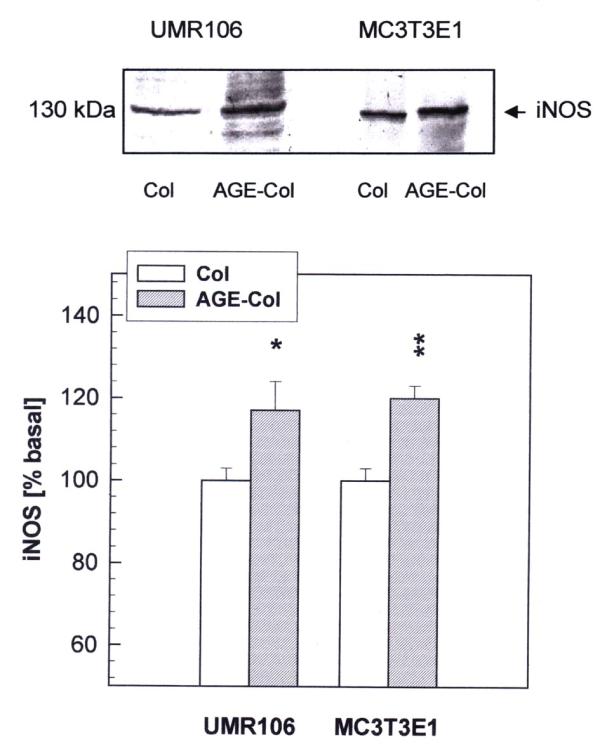
Results: AGE-collagen decreased the adhesion of UMR106 cells, but had no effect on the attachment of MC3T3E1 cells. In the UMR106 cell line, AGE-collagen also inhibited cellular proliferation, spreading and alkaline phosphatase (ALP) activity. In preosteoblastic MC3T3E1 cells (24-hour culture), proliferation and spreading were significantly increased by AGE-collagen. After one week of culture (differentiated MC3T3E1 osteoblasts) AGE-collagen inhibited ALP activity, but had no effect on cell number. In mineralizing MC3T3E1 cells (3-week culture) AGE-collagen induced a decrease in the number of surviving cells and of extracellular nodules of mineralization, without modifying their ALP activity. Intracellular ROS production, measured after a 48-hour culture, was decreased by AGE-collagen in MC3T3E1 cells, but was increased by AGE-collagen in UMR106 cells. After a 24-hour culture, AGE-collagen increased the expression of endothelial and inducible NOS, in both osteoblastic cell lines.
Conclusions: These results suggest that the accumulation of AGE on bone extracellular matrix could regulate the proliferation and differentiation of osteoblastic cells. These effects appear to depend on the stage of osteoblastic development, and possibly involve the modulation of NOS expression and intracellular ROS pathways.
Figures
References
-
- Vlassara H, Bucala R, Striker L. Pathogenic effects of advanced glycosylation: biochemical, biologic and clinical implications for diabetes and ageing. Lab Invest. 1994;70:138–151. - PubMed
-
- Baynes JW, Thorpe SR. Role of oxidative stress in diabetic complications: a new perspective on an old paradigm. Diabetes. 1999;48:1–9. - PubMed
-
- Thornalley PJ. Cell activation by glycated proteins. Age receptors, receptor recognition factors and functional classification of AGEs. Cell Mol Biol (Noisy-le-Grand) 1998;44:1013–1033. - PubMed
-
- Chakravarthy U, Hayes RG, Stitt AW, McAuley E, Archer DB. Constitutive nitric oxide synthase expression in retinal vascular endothelial cells is supported by high glucose and advanced glycation end products. Diabetes. 1998;47:945–952. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
