

Rpm2p: separate domains promote tRNA and Rpm1r maturation in Saccharomyces cerevisiae mitochondria
- PMID: 11522833
- PMCID: PMC55890
- DOI: 10.1093/nar/29.17.3631
Rpm2p: separate domains promote tRNA and Rpm1r maturation in Saccharomyces cerevisiae mitochondria
Abstract
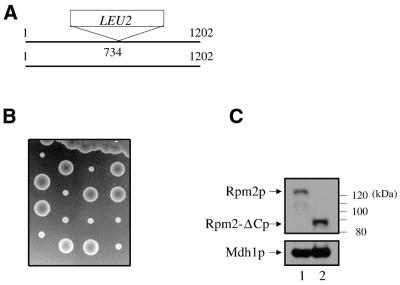
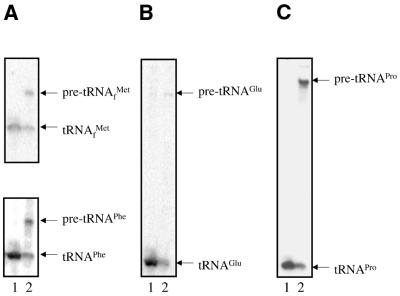
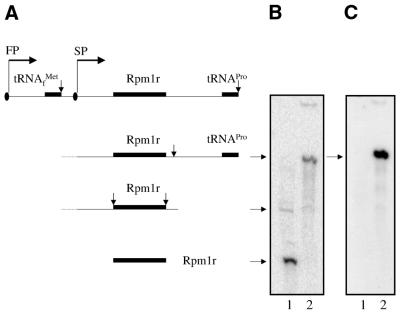
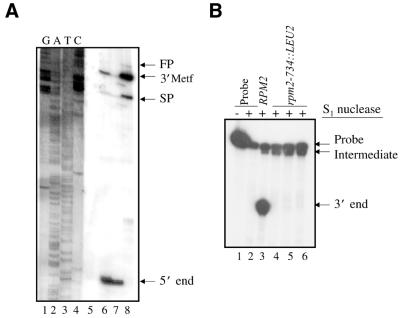
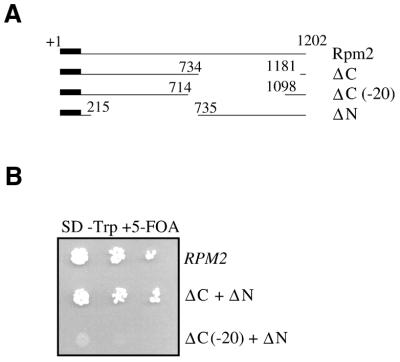
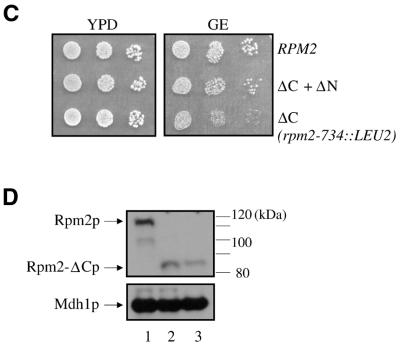
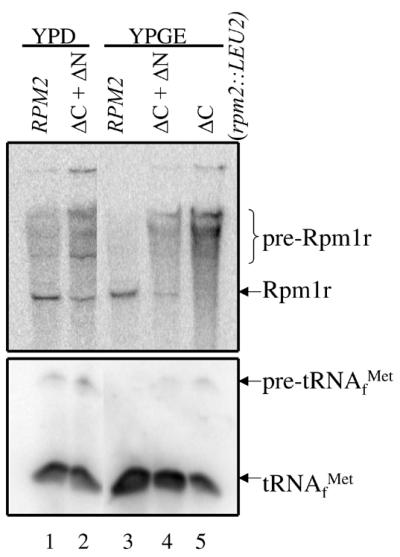
Rpm2p is a protein subunit of yeast mitochondrial RNase P and is also required for the maturation of Rpm1r, the mitochondrially-encoded RNA subunit of the enzyme. Previous work demonstrated that an insertional disruption of RPM2, which produces the C-terminally truncated protein Rpm2-DeltaCp, supports growth on glucose but cells lose some or all of their mitochondrial genome and become petite. These petites, even if they retain the RPM1 locus, lose their ability to process the 5'-ends of mitochondrial tRNA. We report here that if strains containing the truncated RPM2 allele are created and maintained on respiratory carbon sources they have wild-type mitochondrial genomes, and a significant portion of tRNA transcripts are processed. In contrast, precursor Rpm1r transcripts accumulate and mature Rpm1r is not made. These data show that one function of the deleted C-terminal region is in the maturation of Rpm1r, and that this region and mature Rpm1r are not absolutely required for RNase P activity. Finally, we demonstrate that full activity can be restored if the N-terminal and C-terminal domains of Rpm2p are supplied in trans.
Figures
Similar articles
-
Yeast mitochondrial RNase P RNA synthesis is altered in an RNase P protein subunit mutant: insights into the biogenesis of a mitochondrial RNA-processing enzyme.Mol Cell Biol. 1996 Jul;16(7):3429-36. doi: 10.1128/MCB.16.7.3429. Mol Cell Biol. 1996. PMID: 8668158 Free PMC article.
-
Kluyveromyces lactis SEF1 and its Saccharomyces cerevisiae homologue bypass the unknown essential function, but not the mitochondrial RNase P function, of the S. cerevisiae RPM2 gene.Yeast. 1998 Jan 15;14(1):77-87. doi: 10.1002/(SICI)1097-0061(19980115)14:1<77::AID-YEA201>3.0.CO;2-P. Yeast. 1998. PMID: 9483797
-
Successful transformation of yeast mitochondria with RPM1: an approach for in vivo studies of mitochondrial RNase P RNA structure, function and biosynthesis.Nucleic Acids Res. 1995 Mar 11;23(5):856-60. doi: 10.1093/nar/23.5.856. Nucleic Acids Res. 1995. PMID: 7708503 Free PMC article.
-
tRNA transfers to the limelight.Genes Dev. 2003 Jan 15;17(2):162-80. doi: 10.1101/gad.1049103. Genes Dev. 2003. PMID: 12533506 Review. No abstract available.
-
Ribonuclease P: unity and diversity in a tRNA processing ribozyme.Annu Rev Biochem. 1998;67:153-80. doi: 10.1146/annurev.biochem.67.1.153. Annu Rev Biochem. 1998. PMID: 9759486 Review.
Cited by
-
Of P and Z: mitochondrial tRNA processing enzymes.Biochim Biophys Acta. 2012 Sep-Oct;1819(9-10):1017-26. doi: 10.1016/j.bbagrm.2011.11.003. Epub 2011 Nov 23. Biochim Biophys Acta. 2012. PMID: 22137969 Free PMC article. Review.
-
Intersection of RNA processing and the type II fatty acid synthesis pathway in yeast mitochondria.Mol Cell Biol. 2008 Nov;28(21):6646-57. doi: 10.1128/MCB.01162-08. Epub 2008 Sep 8. Mol Cell Biol. 2008. PMID: 18779316 Free PMC article.
-
Eukaryotic ribonuclease P: a plurality of ribonucleoprotein enzymes.Annu Rev Biochem. 2002;71:165-89. doi: 10.1146/annurev.biochem.71.110601.135352. Epub 2001 Nov 9. Annu Rev Biochem. 2002. PMID: 12045094 Free PMC article. Review.
-
Ribonuclease P: the evolution of an ancient RNA enzyme.Crit Rev Biochem Mol Biol. 2006 Mar-Apr;41(2):77-102. doi: 10.1080/10409230600602634. Crit Rev Biochem Mol Biol. 2006. PMID: 16595295 Free PMC article.
-
Rpm2p, a component of yeast mitochondrial RNase P, acts as a transcriptional activator in the nucleus.Mol Cell Biol. 2005 Aug;25(15):6546-58. doi: 10.1128/MCB.25.15.6546-6558.2005. Mol Cell Biol. 2005. PMID: 16024791 Free PMC article.
References
-
- Hopper A.K. and Martin,N.C. (1992) Processing of yeast cytoplasmic and mitochondrial precursor tRNAs. In Jones,E.W., Pringle,J.R. and Broach,J.R. (eds), The Molecular and Cellular Biology of the Yeast Saccharomyces: Gene Expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 99–141.
-
- Frank D.N. and Pace,N.R. (1998) Ribonuclease P: unity and diversity in a tRNA processing enzyme. Annu. Rev. Biochem., 67, 153–180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
