

Functional effects of central core disease mutations in the cytoplasmic region of the skeletal muscle ryanodine receptor
- PMID: 11524458
- PMCID: PMC2229502
- DOI: 10.1085/jgp.118.3.277
Functional effects of central core disease mutations in the cytoplasmic region of the skeletal muscle ryanodine receptor
Abstract
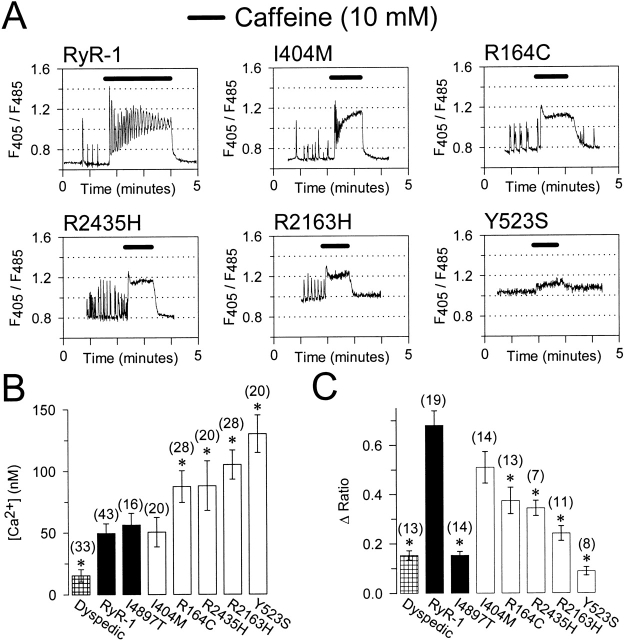
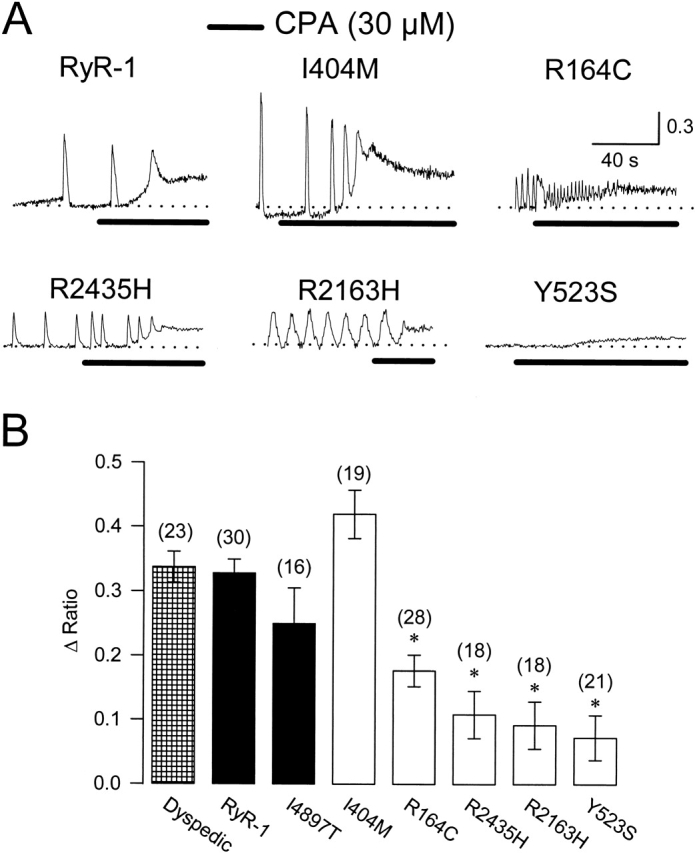
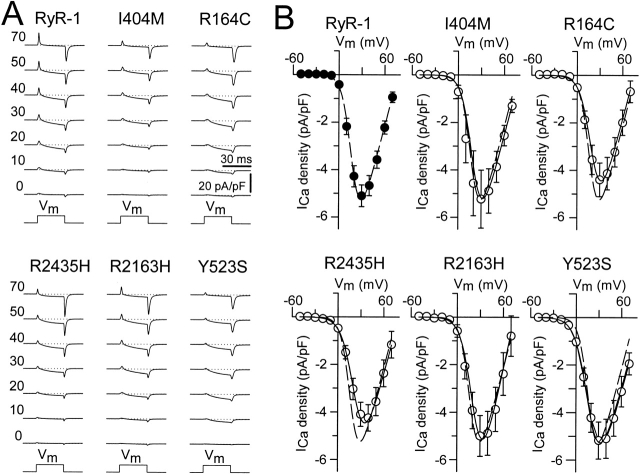
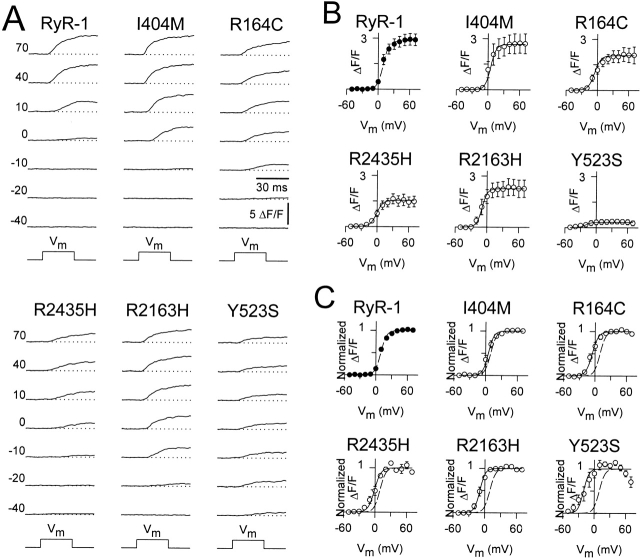
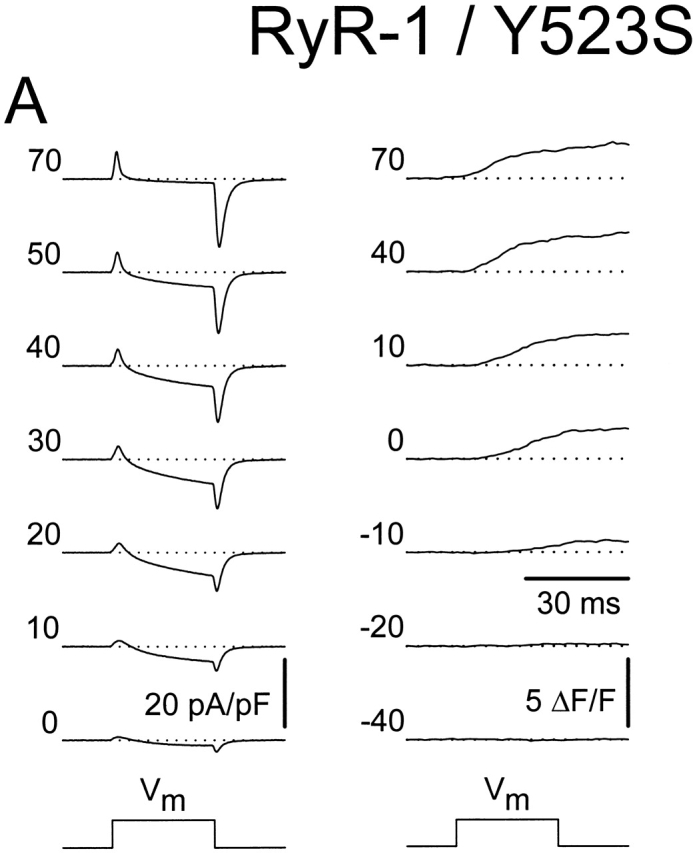
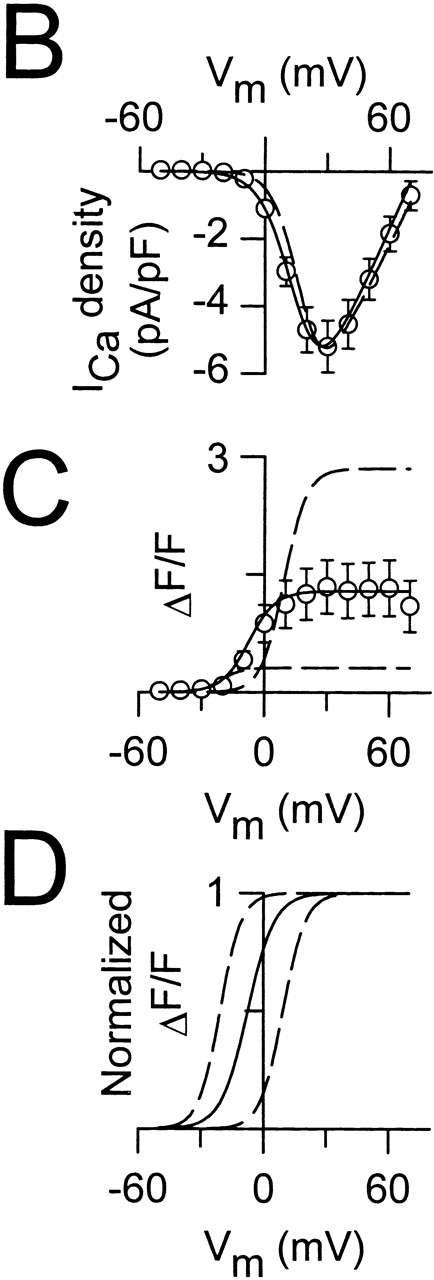
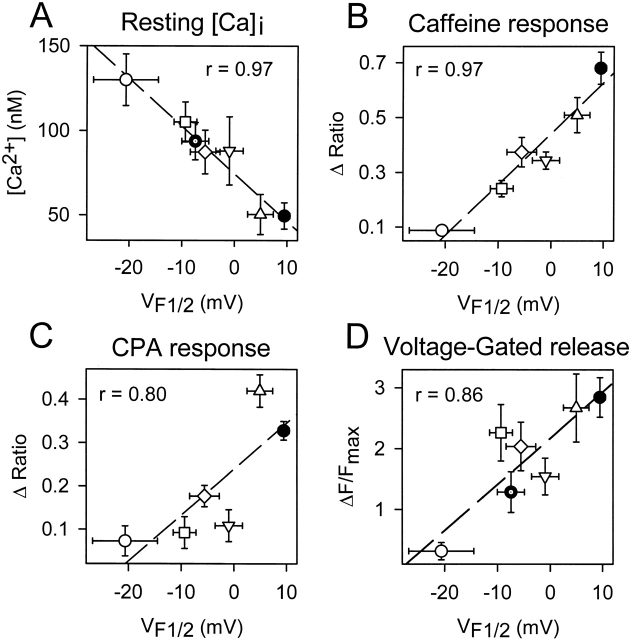
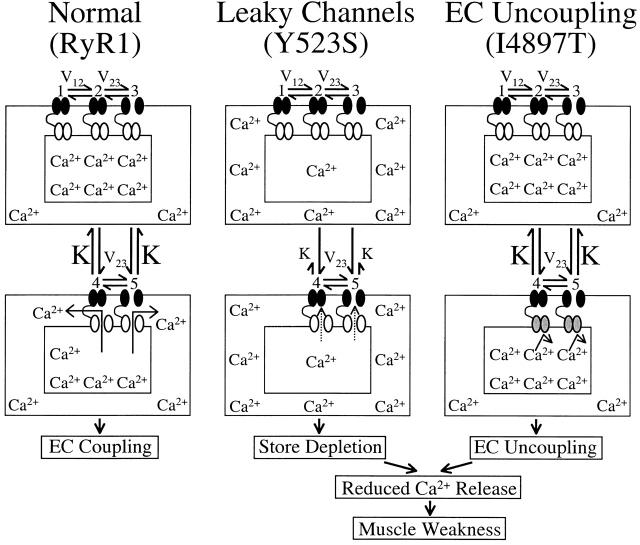
Central core disease (CCD) is a human myopathy that involves a dysregulation in muscle Ca(2)+ homeostasis caused by mutations in the gene encoding the skeletal muscle ryanodine receptor (RyR1), the protein that comprises the calcium release channel of the SR. Although genetic studies have clearly demonstrated linkage between mutations in RyR1 and CCD, the impact of these mutations on release channel function and excitation-contraction coupling in skeletal muscle is unknown. Toward this goal, we have engineered the different CCD mutations found in the NH(2)-terminal region of RyR1 into a rabbit RyR1 cDNA (R164C, I404M, Y523S, R2163H, and R2435H) and characterized the functional effects of these mutations after expression in myotubes derived from RyR1-knockout (dyspedic) mice. Resting Ca(2)+ levels were elevated in dyspedic myotubes expressing four of these mutants (Y523S > R2163H > R2435H R164C > I404M RyR1). A similar rank order was also found for the degree of SR Ca(2)+ depletion assessed using maximal concentrations of caffeine (10 mM) or cyclopiazonic acid (CPA, 30 microM). Although all of the CCD mutants fully restored L-current density, voltage-gated SR Ca(2)+ release was smaller and activated at more negative potentials for myotubes expressing the NH(2)-terminal CCD mutations. The shift in the voltage dependence of SR Ca(2)+ release correlated strongly with changes in resting Ca(2)+, SR Ca(2)+ store depletion, and peak voltage-gated release, indicating that increased release channel activity at negative membrane potentials promotes SR Ca(2)+ leak. Coexpression of wild-type and Y523S RyR1 proteins in dyspedic myotubes resulted in release channels that exhibited an intermediate degree of SR Ca(2)+ leak. These results demonstrate that the NH(2)-terminal CCD mutants enhance release channel sensitivity to activation by voltage in a manner that leads to increased SR Ca(2)+ leak, store depletion, and a reduction in voltage-gated Ca(2)+ release. Two fundamentally distinct cellular mechanisms (leaky channels and EC uncoupling) are proposed to explain how altered release channel function caused by different mutations in RyR1 could result in muscle weakness in CCD.
Figures
References
-
- Avila G., O'Connell K.M.S., Groom L., Dirksen R.T. Ca2+ release through ryanodine receptors regulates skeletal muscle L-type Ca2+ channel expression J. Biol. Chem. 276 2001. 17732 17738b - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
