

KcsA: it's a potassium channel
- PMID: 11524460
- PMCID: PMC2229506
- DOI: 10.1085/jgp.118.3.303
KcsA: it's a potassium channel
Abstract
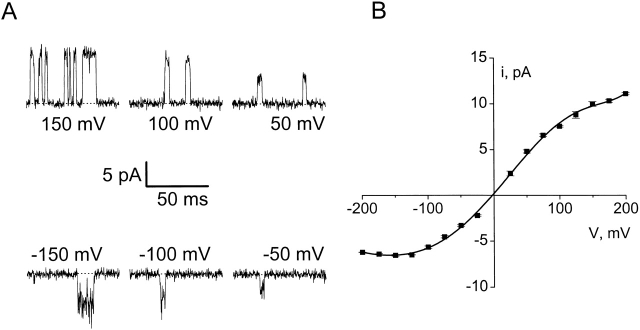
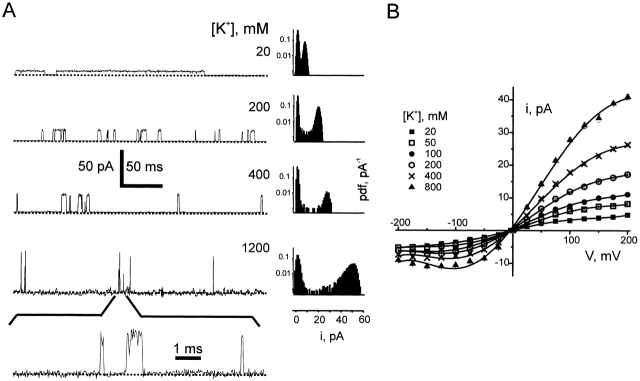
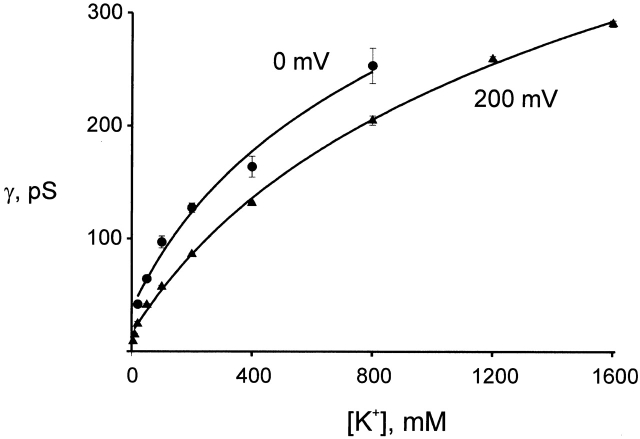
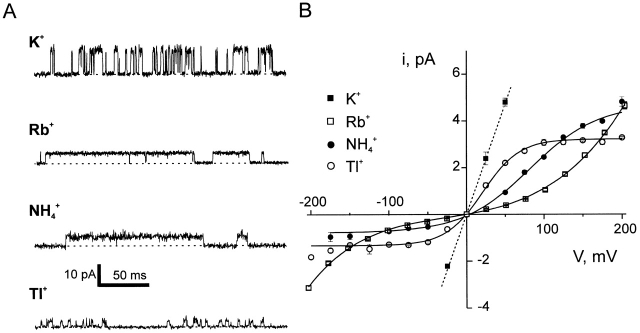
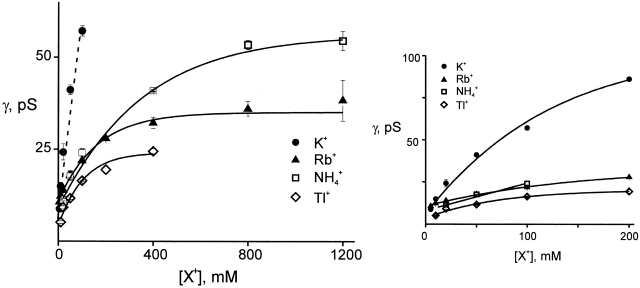
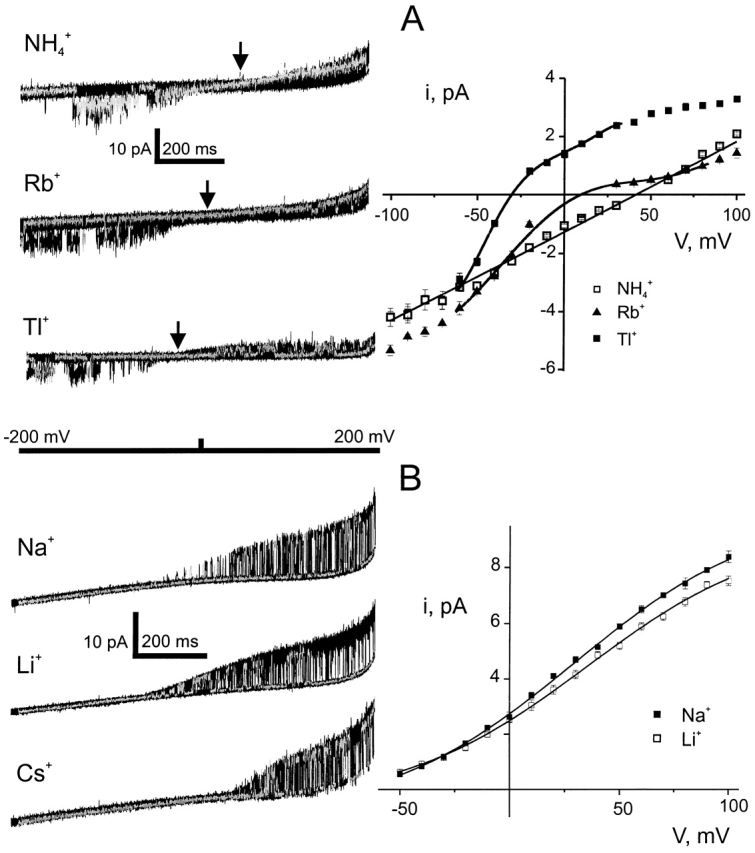
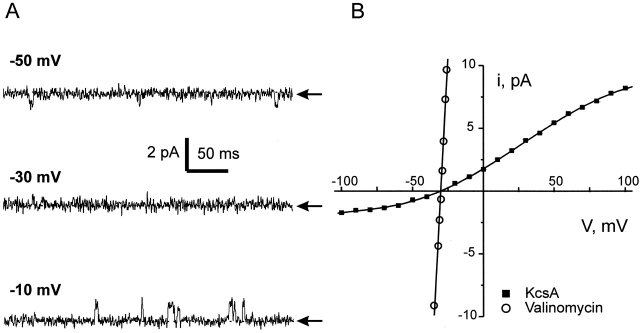
Ion conduction and selectivity properties of KcsA, a bacterial ion channel of known structure, were studied in a planar lipid bilayer system at the single-channel level. Selectivity sequences for permeant ions were determined by symmetrical solution conductance (K(+) > Rb(+), NH(4)(+), Tl(+) >> Cs(+), Na(+), Li(+)) and by reversal potentials under bi-ionic or mixed-ion conditions (Tl(+) > K(+) > Rb(+) > NH(4)(+) >> Na(+), Li(+)). Determination of reversal potentials with submillivolt accuracy shows that K(+) is over 150-fold more permeant than Na(+). Variation of conductance with concentration under symmetrical salt conditions is complex, with at least two ion-binding processes revealing themselves: a high affinity process below 20 mM and a low affinity process over the range 100-1,000 mM. These properties are analogous to those seen in many eukaryotic K(+) channels, and they establish KcsA as a faithful structural model for ion permeation in eukaryotic K(+) channels.
Figures
References
-
- Abramson J.J., Shamoo A.E. Anionic detergents as divalent cation ionophores across black lipid membranes. J. Membr. Biol. 1979;50:241–255. - PubMed
