

Autophagy in yeast: mechanistic insights and physiological function
- PMID: 11528006
- PMCID: PMC99037
- DOI: 10.1128/MMBR.65.3.463-479.2001
Autophagy in yeast: mechanistic insights and physiological function
Abstract
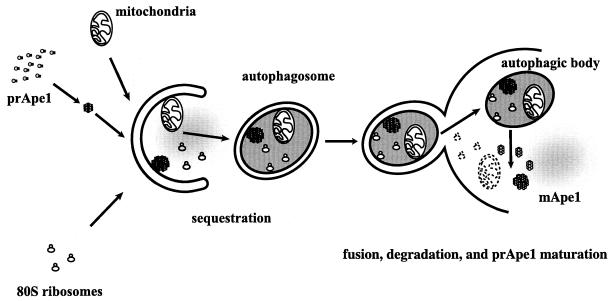
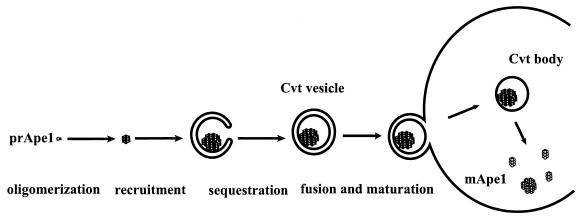
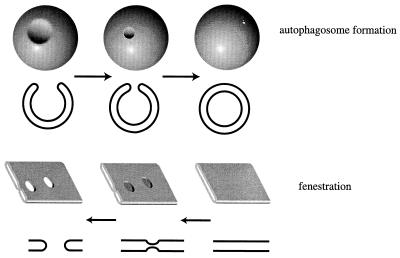
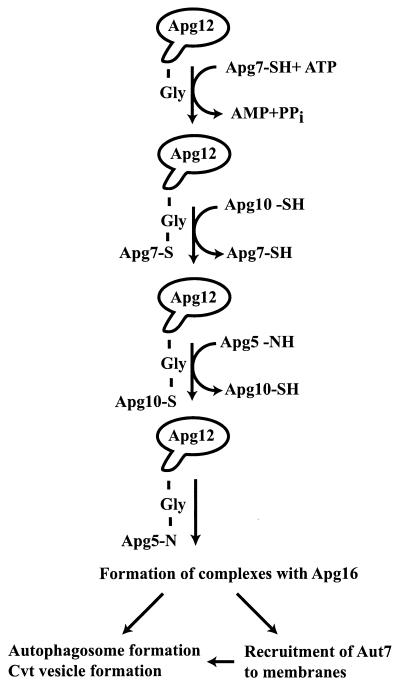
Unicellular eukaryotic organisms must be capable of rapid adaptation to changing environments. While such changes do not normally occur in the tissues of multicellular organisms, developmental and pathological changes in the environment of cells often require adaptation mechanisms not dissimilar from those found in simpler cells. Autophagy is a catabolic membrane-trafficking phenomenon that occurs in response to dramatic changes in the nutrients available to yeast cells, for example during starvation or after challenge with rapamycin, a macrolide antibiotic whose effects mimic starvation. Autophagy also occurs in animal cells that are serum starved or challenged with specific hormonal stimuli. In macroautophagy, the form of autophagy commonly observed, cytoplasmic material is sequestered in double-membrane vesicles called autophagosomes and is then delivered to a lytic compartment such as the yeast vacuole or mammalian lysosome. In this fashion, autophagy allows the degradation and recycling of a wide spectrum of biological macromolecules. While autophagy is induced only under specific conditions, salient mechanistic aspects of autophagy are functional in a constitutive fashion. In Saccharomyces cerevisiae, induction of autophagy subverts a constitutive membrane-trafficking mechanism called the cytoplasm-to-vacuole targeting pathway from a specific mode, in which it carries the resident vacuolar hydrolase, aminopeptidase I, to a nonspecific bulk mode in which significant amounts of cytoplasmic material are also sequestered and recycled in the vacuole. The general aim of this review is to focus on insights gained into the mechanism of autophagy in yeast and also to review our understanding of the physiological significance of autophagy in both yeast and higher organisms.
Figures
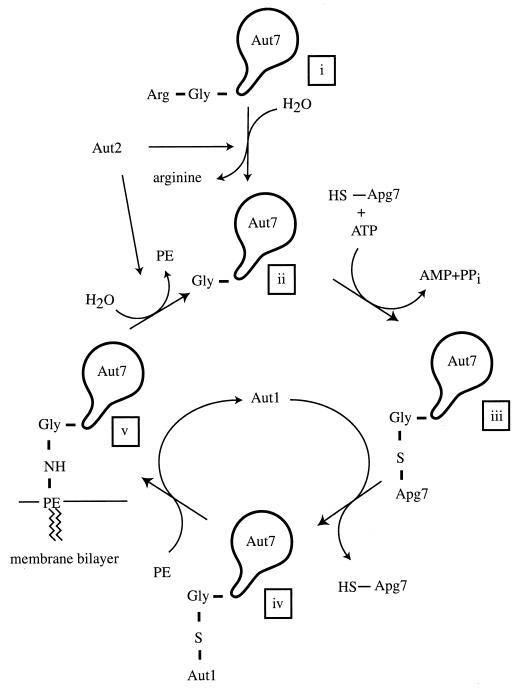
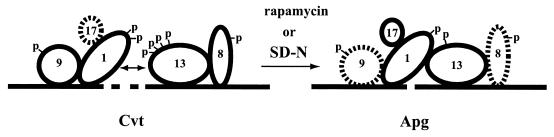
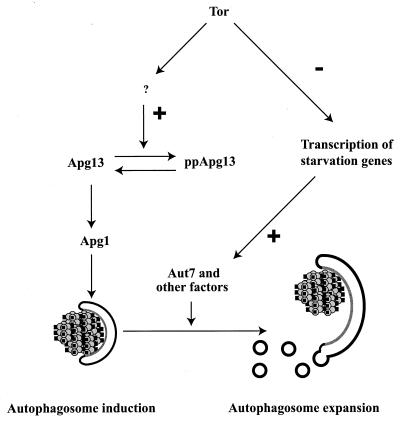
References
-
- Abeliovich H, Grote E, Novick P, Ferro-Novick S. Tlg2p, a yeast syntaxin homolog that resides on the Golgi and endocytic structures. J Biol Chem. 1998;273:11719–11727. - PubMed
-
- Anglade P, Vyas S, Javoy-Agid F, Herrero M T, Michel P P, Marquez J, Mouatt-Progent A, Ruberg M, Hirsch E C, Agid Y. Apoptosis and autophagy in nigral neurons of patients with Parkinson's disease. Histol Histopathol. 1997;12:25–31. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
