

Isolated lymphatic endothelial cells transduce growth, survival and migratory signals via the VEGF-C/D receptor VEGFR-3
- PMID: 11532940
- PMCID: PMC125596
- DOI: 10.1093/emboj/20.17.4762
Isolated lymphatic endothelial cells transduce growth, survival and migratory signals via the VEGF-C/D receptor VEGFR-3
Abstract
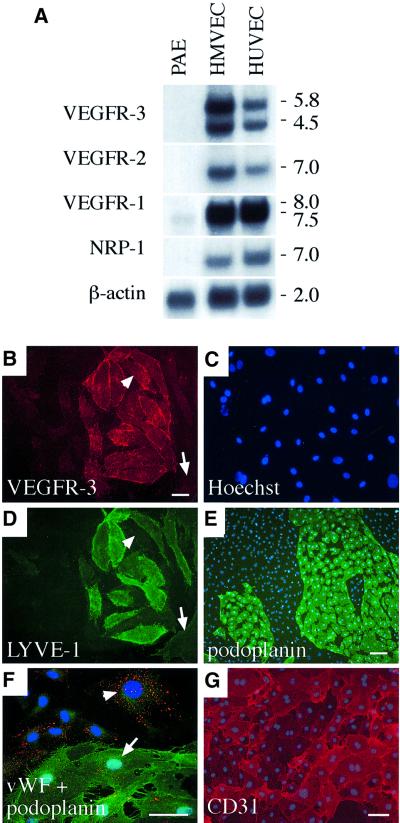
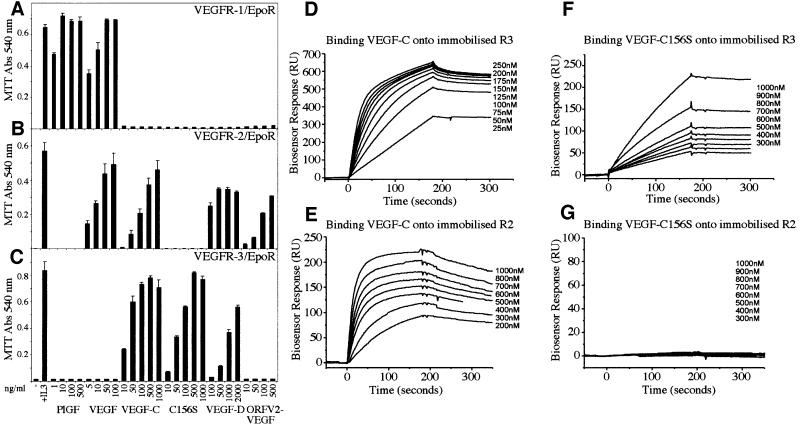
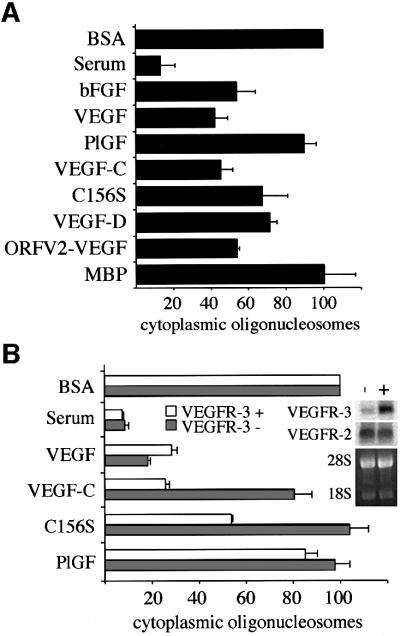
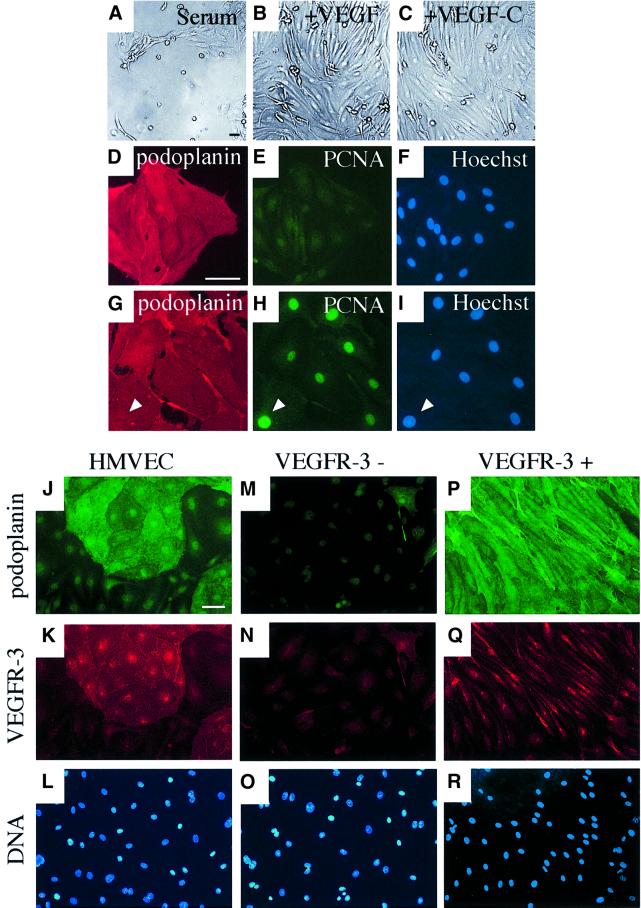
Vascular endothelial growth factor receptor-3 (VEGFR-3/Flt4) binds two known members of the VEGF ligand family, VEGF-C and VEGF-D, and has a critical function in the remodelling of the primary capillary vasculature of midgestation embryos. Later during development, VEGFR-3 regulates the growth and maintenance of the lymphatic vessels. In the present study, we have isolated and cultured stable lineages of blood vascular and lymphatic endothelial cells from human primary microvascular endothelium by using antibodies against the extracellular domain of VEGFR-3. We show that VEGFR-3 stimulation alone protects the lymphatic endothelial cells from serum deprivation-induced apoptosis and induces their growth and migration. At least some of these signals are transduced via a protein kinase C-dependent activation of the p42/p44 MAPK signalling cascade and via a wortmannin-sensitive induction of Akt phosphorylation. These results define the critical role of VEGF-C/VEGFR-3 signalling in the growth and survival of lymphatic endothelial cells. The culture of isolated lymphatic endothelial cells should now allow further studies of the molecular properties of these cells.
Figures
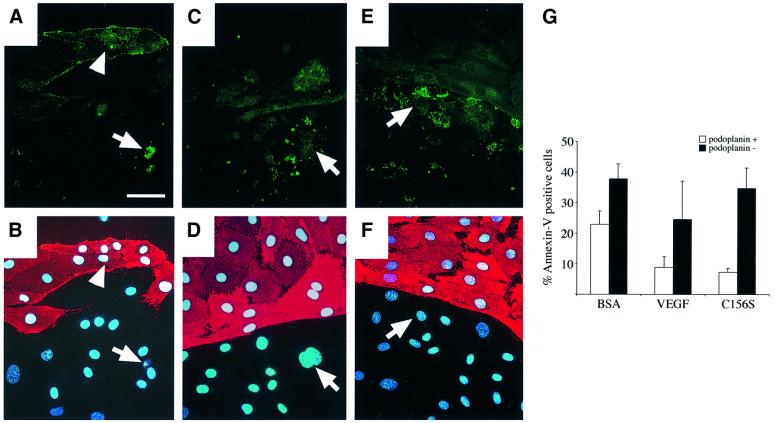
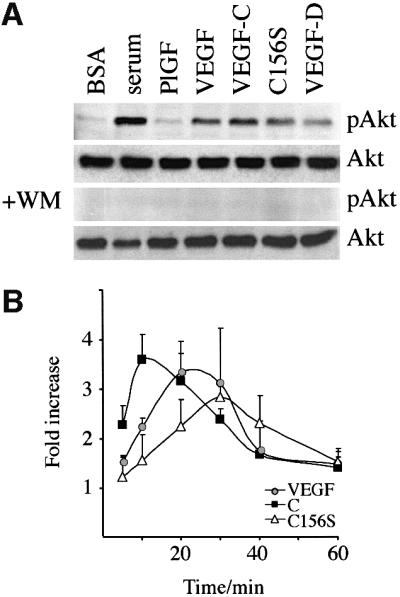
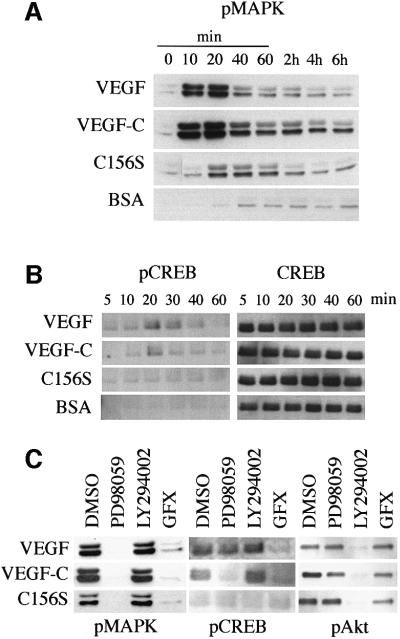
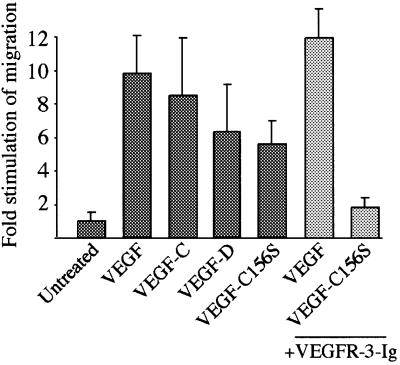
References
-
- Achen M.G. et al. (2000) Monoclonal antibodies to vascular endothelial growth factor-D block its interactions with both VEGF receptor-2 and VEGF receptor-3. Eur. J. Biochem., 267, 2505–2515. - PubMed
-
- Aiello L.P., Pierce,E.A., Foley,E.D., Takagi,H., Chen,H., Riddle,L., Ferrara,N., King,G.L. and Smith,L.E. (1995) Suppression of retinal neovascularization in vivo by inhibition of vascular endothelial growth factor (VEGF) using soluble VEGF-receptor chimeric proteins. Proc. Natl Acad. Sci. USA, 92, 10457–10461. - PMC - PubMed
-
- Baldwin M.E. et al. (2001) The specificity of receptor binding by vascular endothelial growth factor-D is different in mouse and man. J. Biol. Chem., 276, 19166–19171. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
