

The exon-exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay
- PMID: 11532962
- PMCID: PMC125616
- DOI: 10.1093/emboj/20.17.4987
The exon-exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay
Abstract
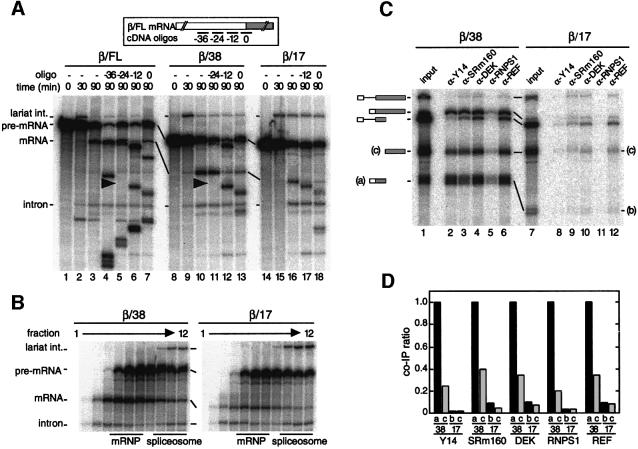
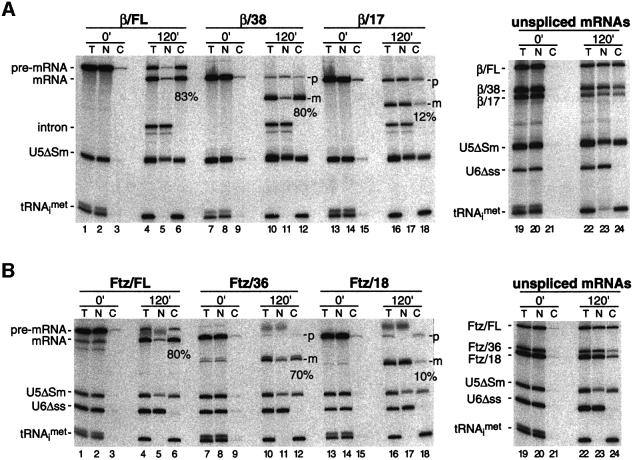
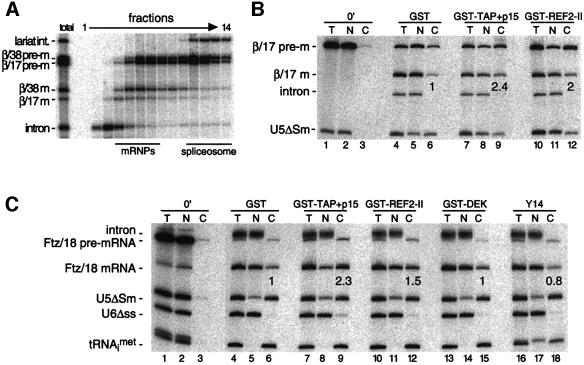
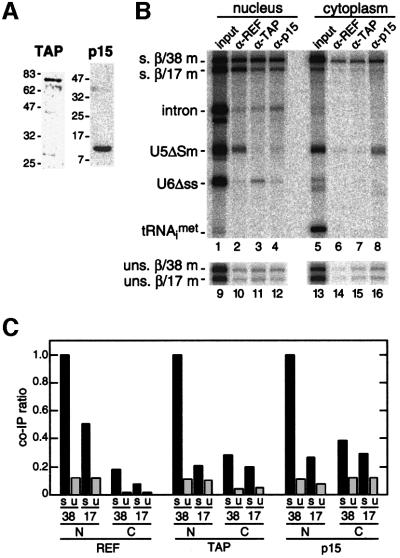
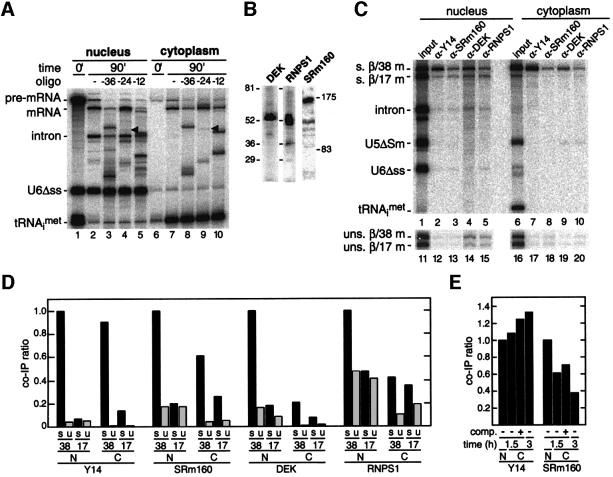
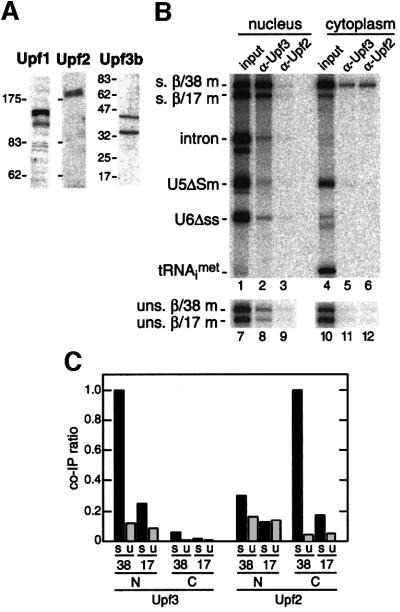
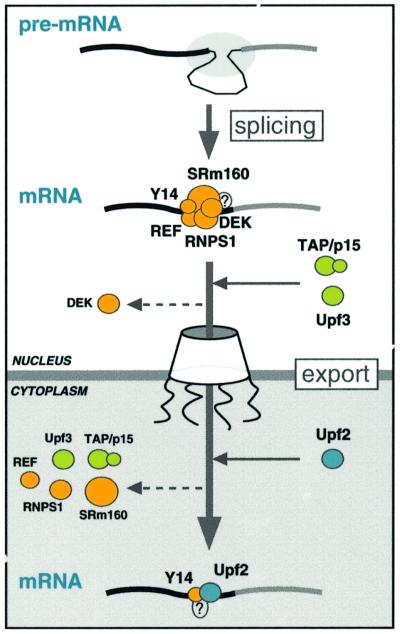
We recently reported that spliceosomes alter messenger ribonucleoprotein particle (mRNP) composition by depositing several proteins 20-24 nucleotides upstream of mRNA exon-exon junctions. When assembled in vitro, this so-called 'exon-exon junction complex' (EJC) contains at least five proteins: SRm160, DEK, RNPS1, Y14 and REF. To better investigate its functional attributes, we now describe a method for generating spliced mRNAs both in vitro and in vivo that either do or do not carry the EJC. Analysis of these mRNAs in Xenopus laevis oocytes revealed that this complex is the species responsible for enhancing nucleocytoplasmic export of spliced mRNAs. It does so by providing a strong binding site for the mRNA export factors REF and TAP/p15. Moreover, by serving as an anchoring point for the factors Upf2 and Upf3, the EJC provides a direct link between splicing and nonsense-mediated mRNA decay. Finally, we show that the composition of the EJC is dynamic in vivo and is subject to significant evolution upon mRNA export to the cytoplasm.
Figures
References
-
- Braun I.C., Herold,A., Rode,M., Conti,E. and Izaurralde,E. (2001) Overexpression of TAP/p15 heterodimers bypasses nuclear retention and stimulates nuclear mRNA export. J. Biol. Chem., 276, 20536–20543. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
