

Adeno-associated virus type 2-mediated gene transfer: role of cellular FKBP52 protein in transgene expression
- PMID: 11533160
- PMCID: PMC114465
- DOI: 10.1128/JVI.75.19.8968-8976.2001
Adeno-associated virus type 2-mediated gene transfer: role of cellular FKBP52 protein in transgene expression
Abstract
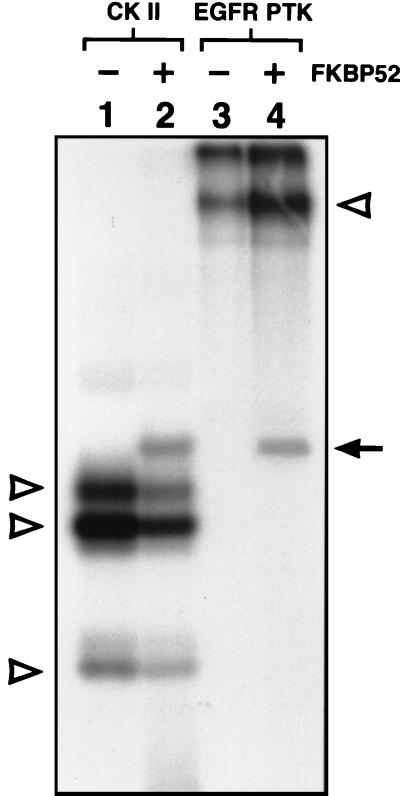
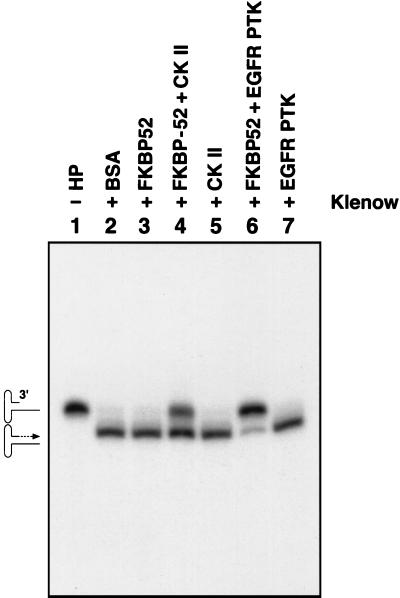
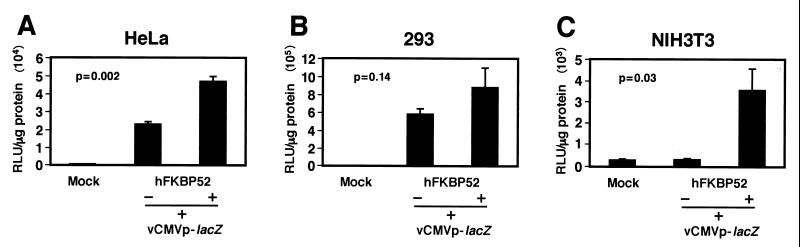
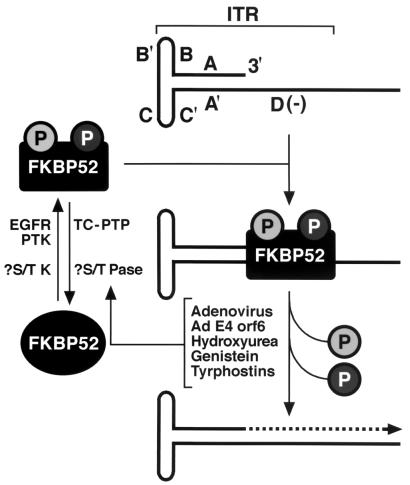
Although adeno-associated virus type 2 (AAV) has gained attention as a potentially useful vector for human gene therapy, the transduction efficiencies of AAV vectors vary greatly in different cells and tissues in vitro and in vivo. We have documented that a cellular tyrosine phosphoprotein, designated the single-stranded D-sequence-binding protein (ssD-BP), plays a crucial role in AAV-mediated transgene expression (K. Y. Qing, X.-S. Wang, D. M. Kube, S. Ponnazhagan, A. Bajpai, and A. Srivastava, Proc. Natl. Acad. Sci. USA 94:10879-10884, 1997). We have documented a strong correlation between the phosphorylation state of ssD-BP and AAV transduction efficiency in vitro as well as in vivo (K. Y. Qing, B. Khuntrirat, C. Mah, D. M. Kube, X.-S. Wang, S. Ponnazhagan, S. Z. Zhou, V. J. Dwarki, M. C. Yoder, and A. Srivastava, J. Virol. 72:1593-1599, 1998). We have also established that the ssD-BP is phosphorylated by epidermal growth factor receptor protein tyrosine kinase and that the tyrosine-phosphorylated form, but not the dephosphorylated form, of ssD-BP prevents AAV second-strand DNA synthesis and, consequently, results in a significant inhibition of AAV-mediated transgene expression (C. Mah, K. Y. Qing, B. Khuntrirat, S. Ponnazhagan, X.-S. Wang, D. M. Kube, M. C. Yoder, and A. Srivastava, J. Virol. 72:9835-9841, 1998). Here, we report that a partial amino acid sequence of ssD-BP purified from HeLa cells is identical to a portion of a cellular protein that binds the immunosuppressant drug FK506, termed the FK506-binding protein 52 (FKBP52). FKBP52 was purified by using a prokaryotic expression plasmid containing the human cDNA. The purified protein could be phosphorylated at both tyrosine and serine or threonine residues, and only the phosphorylated forms of FKBP52 were shown to interact with the AAV single-stranded D-sequence probe. Furthermore, in in vitro DNA replication assays, tyrosine-phosphorylated FKBP52 inhibited AAV second-strand DNA synthesis by greater than 90%. Serine- or threonine-phosphorylated FKBP52 caused approximately 40% inhibition, whereas dephosphorylated FKBP52 had no effect on AAV second-strand DNA synthesis. Deliberate overexpression of FKBP52 effectively reduced the extent of tyrosine phosphorylation of the protein, resulting in a significant increase in AAV-mediated transgene expression in human and murine cell lines. These studies corroborate the idea that the phosphorylation status of the cellular FKBP52 protein correlates strongly with AAV transduction efficiency, which may have important implications for the optimal use of AAV vectors in human gene therapy.
Figures





References
-
- Berns K I, Giraud C. Biology of adeno-associated virus. Curr Top Microbiol Immunol. 1996;218:1–23. - PubMed
-
- Carter B J, Flotte T R. Development of adeno-associated virus vectors for gene therapy of cystic fibrosis. Curr Top Microbiol Immunol. 1996;218:119–144. - PubMed
-
- Chatterjee S, Lu D, Podsakoff G, Wong K K., Jr Strategies for efficient gene transfer into hematopoietic cells: the use of adeno-associated virus vectors in gene therapy. Ann N Y Acad Sci. 1995;770:79–90. - PubMed
-
- Cybulsky A V, Goodyer P R, McTavish A J. Epidermal growth factor receptor activation in developing rat kidney. Am J Physiol. 1994;267:F428–F436. - PubMed
-
- Czar M J, Lyons R H, Welsh M J, Renoir J-M, Pratt W B. Evidence that the FK506-binding protein 56 is required for trafficking of the glucocorticoid receptor from the cytoplasm to the nucleus. Mol Endocrinol. 1995;9:1549–1560. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
