

Molecular basis for the relative substrate specificity of human immunodeficiency virus type 1 and feline immunodeficiency virus proteases
- PMID: 11533208
- PMCID: PMC114513
- DOI: 10.1128/JVI.75.19.9458-9469.2001
Molecular basis for the relative substrate specificity of human immunodeficiency virus type 1 and feline immunodeficiency virus proteases
Abstract
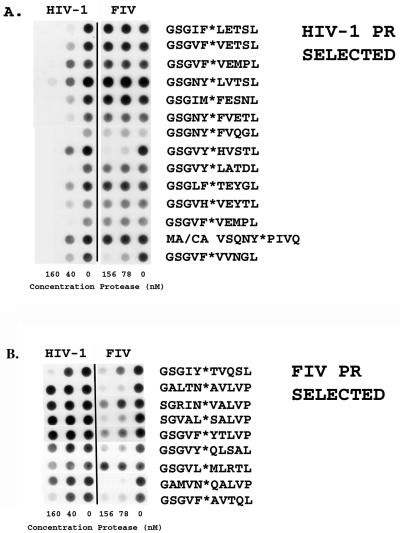
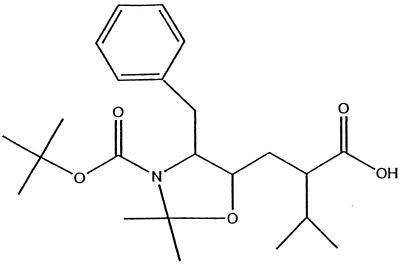
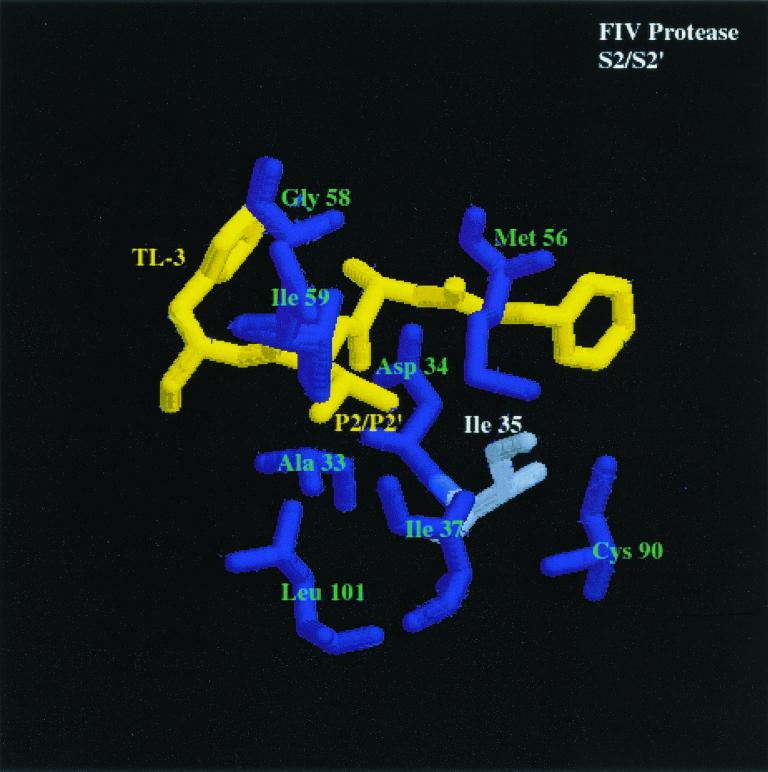
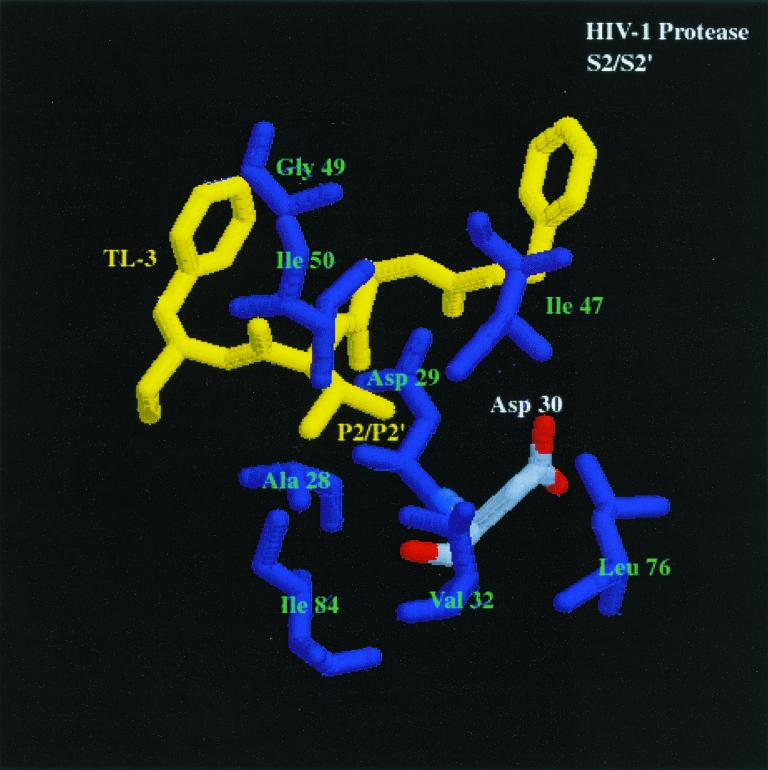
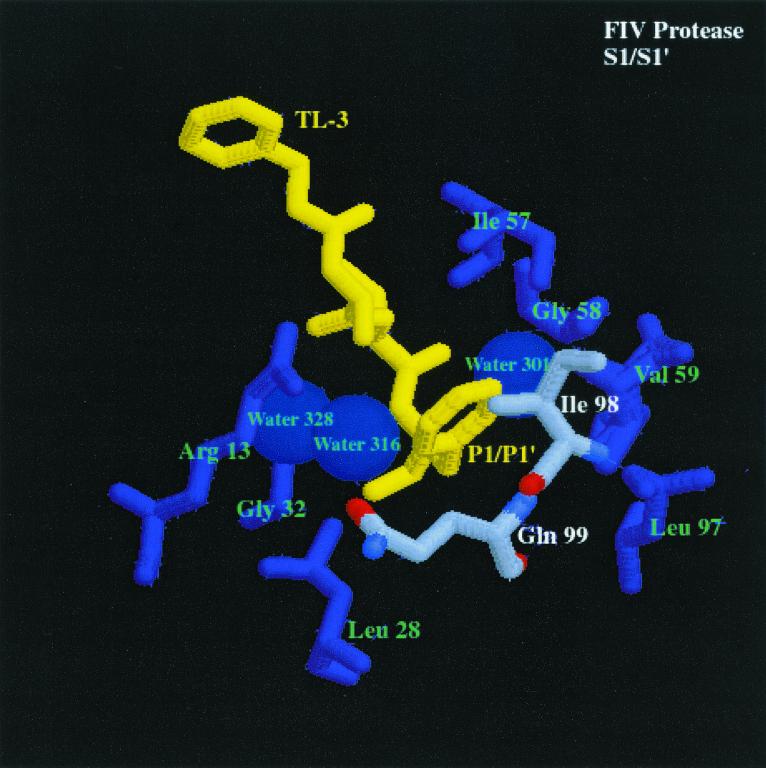
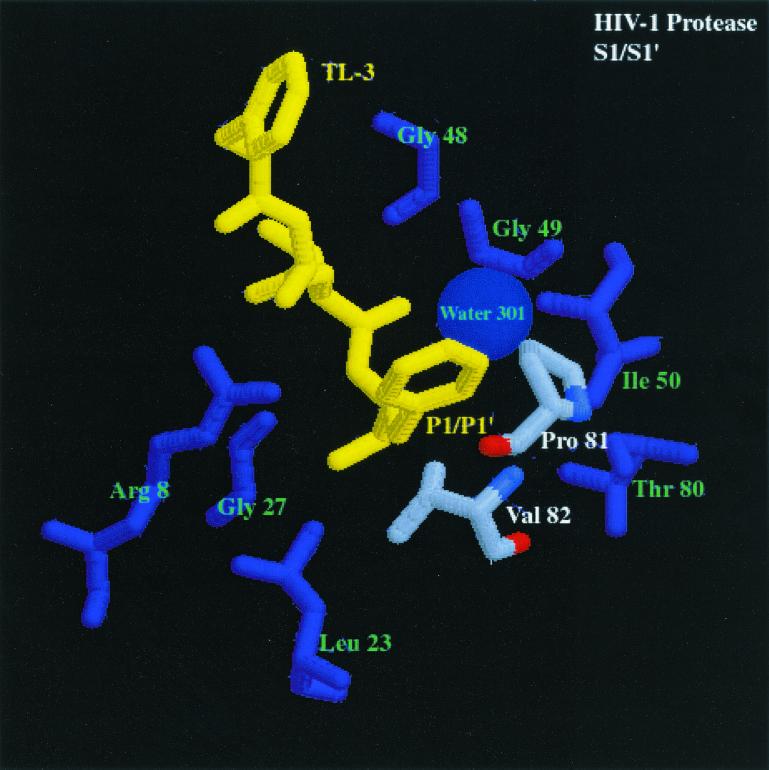
We have used a random hexamer phage library to delineate similarities and differences between the substrate specificities of human immunodeficiency virus type 1 (HIV-1) and feline immunodeficiency virus (FIV) proteases (PRs). Peptide sequences were identified that were specifically cleaved by each protease, as well as sequences cleaved equally well by both enzymes. Based on amino acid distinctions within the P3-P3' region of substrates that appeared to correlate with these cleavage specificities, we prepared a series of synthetic peptides within the framework of a peptide sequence cleaved with essentially the same efficiency by both HIV-1 and FIV PRs, Ac-KSGVF/VVNGLVK-NH(2) (arrow denotes cleavage site). We used the resultant peptide set to assess the influence of specific amino acid substitutions on the cleavage characteristics of the two proteases. The findings show that when Asn is substituted for Val at the P2 position, HIV-1 PR cleaves the substrate at a much greater rate than does FIV PR. Likewise, Glu or Gln substituted for Val at the P2' position also yields peptides specifically susceptible to HIV-1 PR. In contrast, when Ser is substituted for Val at P1', FIV PR cleaves the substrate at a much higher rate than does HIV-1 PR. In addition, Asn or Gln at the P1 position, in combination with an appropriate P3 amino acid, Arg, also strongly favors cleavage by FIV PR over HIV PR. Structural analysis identified several protease residues likely to dictate the observed specificity differences. Interestingly, HIV PR Asp30 (Ile-35 in FIV PR), which influences specificity at the S2 and S2' subsites, and HIV-1 PR Pro-81 and Val-82 (Ile-98 and Gln-99 in FIV PR), which influence specificity at the S1 and S1' subsites, are residues which are often involved in development of drug resistance in HIV-1 protease. The peptide substrate KSGVF/VVNGK, cleaved by both PRs, was used as a template for the design of a reduced amide inhibitor, Ac-GSGVF Psi(CH(2)NH)VVNGL-NH(2.) This compound inhibited both FIV and HIV-1 PRs with approximately equal efficiency. These findings establish a molecular basis for distinctions in substrate specificity between human and feline lentivirus PRs and offer a framework for development of efficient broad-based inhibitors.
Figures
References
-
- Alewood P, Alewood D, Miranda L, Love S, Meutermans W, Wilson D. Rapid in situ neutralization protocols for Boc and Fmoc solid-phase chemistries. Methods Enzymol. 1997;289:14–29. - PubMed
-
- Beck Z Q, Hervio L, Dawson P E, Elder J H, Madison E L. Identification of efficiently cleaved substrates for HIV-1 protease using a phage display library and use in inhibitor development. Virology. 2000;274:391–401. - PubMed
-
- Billich A, Winkler G. Analysis of subsite preferences of HIV-1 proteinase using MA/CA junction peptides substituted at the P3–P1′ positions. Arch Biochem Biophys. 1991;290:186–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
