

Creation of a pluripotent ubiquitin-conjugating enzyme
- PMID: 11533242
- PMCID: PMC99800
- DOI: 10.1128/MCB.21.19.6537-6548.2001
Creation of a pluripotent ubiquitin-conjugating enzyme
Abstract
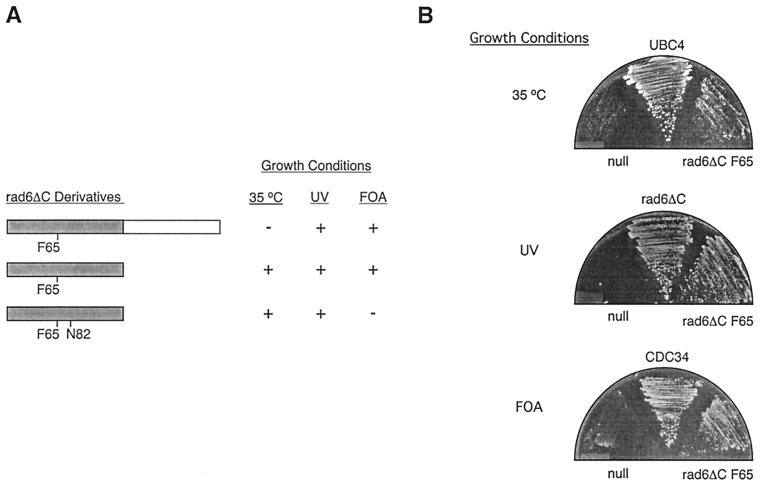
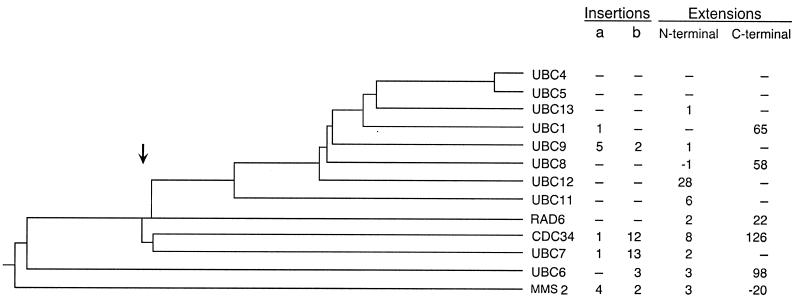
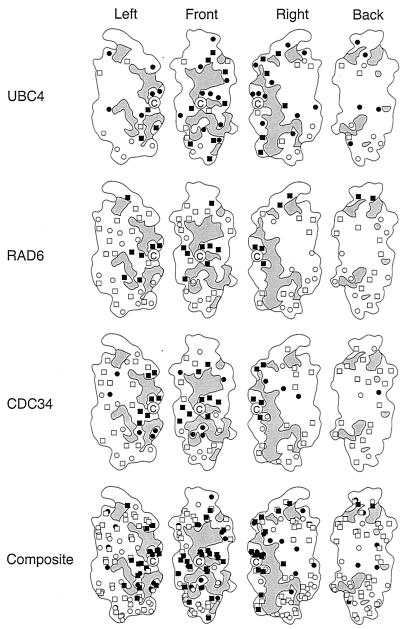
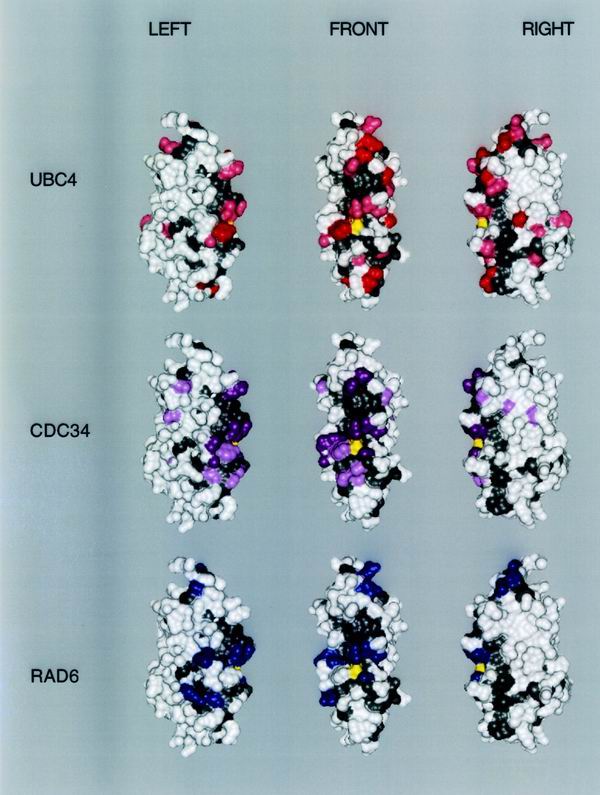
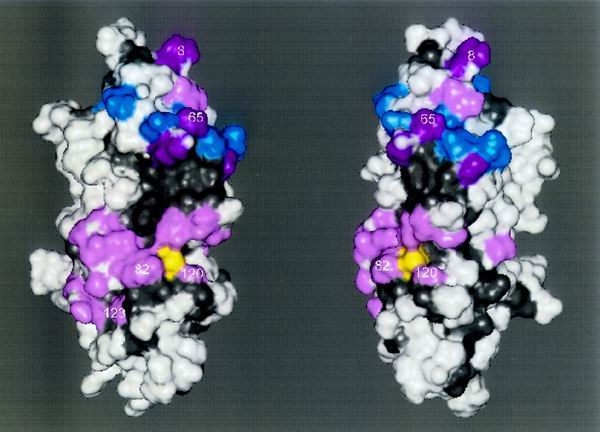
We describe the creation of a pluripotent ubiquitin-conjugating enzyme (E2) generated through a single amino acid substitution within the catalytic domain of RAD6 (UBC2). This RAD6 derivative carries out the stress-related function of UBC4 and the cell cycle function of CDC34 while maintaining its own DNA repair function. Furthermore, it carries out CDC34's function in the absence of the CDC34 carboxy-terminal extension. By using sequence and structural comparisons, the residues that define the unique functions of these three E2s were found on the E2 catalytic face partitioned to either side by a conserved divide. One of these patches corresponds to a binding site for both HECT and RING domain proteins, suggesting that a single substitution in the catalytic domain of RAD6 confers upon it the ability to interact with multiple ubiquitin protein ligases (E3s). Other amino acid substitutions made within the catalytic domain of RAD6 either caused loss of its DNA repair function or modified its ability to carry out multiple E2 functions. These observations suggest that while HECT and RING domain binding may generally be localized to a specific patch on the E2 surface, other regions of the functional E2 face also play a role in specificity. Finally, these data also indicate that RAD6 uses a different functional region than either UBC4 or CDC34, allowing it to acquire the functions of these E2s while maintaining its own. The pluripotent RAD6 derivative, coupled with sequence, structural, and phylogenetic data, suggests that E2s have diverged from a common multifunctional progenitor.
Figures

References
-
- Chen P, Johnson P, Sommer T, Jentsch S, Hochstrasser M. Multiple ubiquitin-conjugating enzymes participate in the in vivo degradation of the yeast MATα2 repressor. Cell. 1993;74:357–369. - PubMed
-
- Cook W J, Jeffery L C, Sullivan M L, Vierstra R D. Three-dimensional structure of a ubiquitin-conjugating enzyme E2. J Biol Chem. 1992;267:15116–15121. - PubMed
-
- Cook W J, Jeffery L C, Xu Y, Chau V. Tertiary structures of class I ubiquitin-conjugating enzymes are highly conserved: crystal structure of yeast UBC4. Biochemistry. 1993;32:13809–13817. - PubMed
-
- Cook W J, Martin P D, Edwards B F, Yamazaki R K, Chau V. Crystal structure of a class I ubiquitin conjugating enzyme (UBC7) from Saccharomyces cerevisiae at 2.9 angstroms resolution. Biochemistry. 1997;36:1621–1627. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials