

The ornithine decarboxylase gene is essential for cell survival during early murine development
- PMID: 11533243
- PMCID: PMC99801
- DOI: 10.1128/MCB.21.19.6549-6558.2001
The ornithine decarboxylase gene is essential for cell survival during early murine development
Abstract
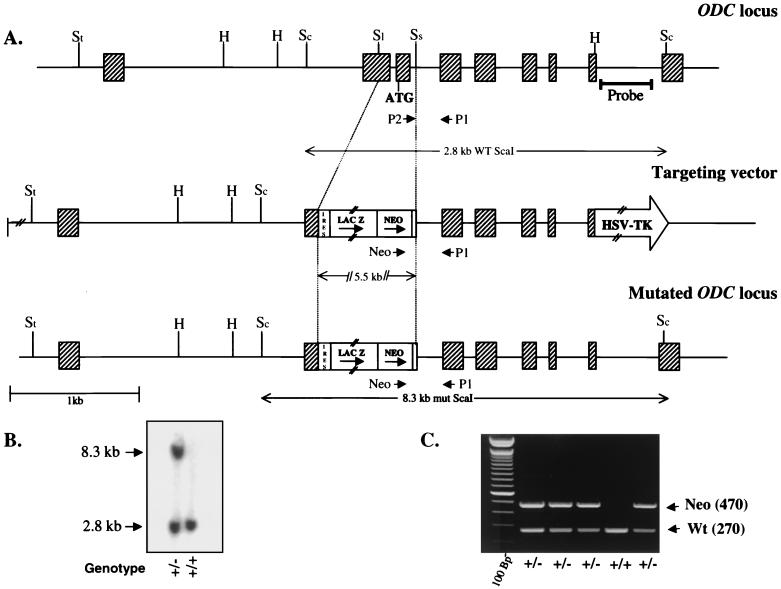
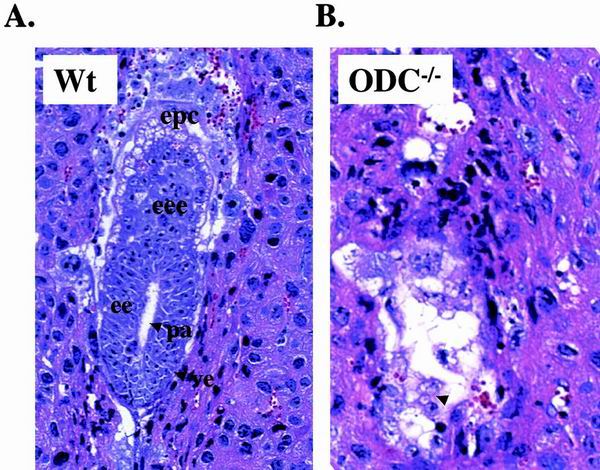
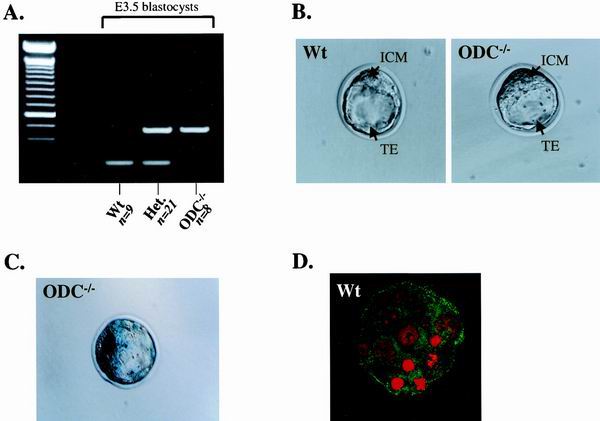
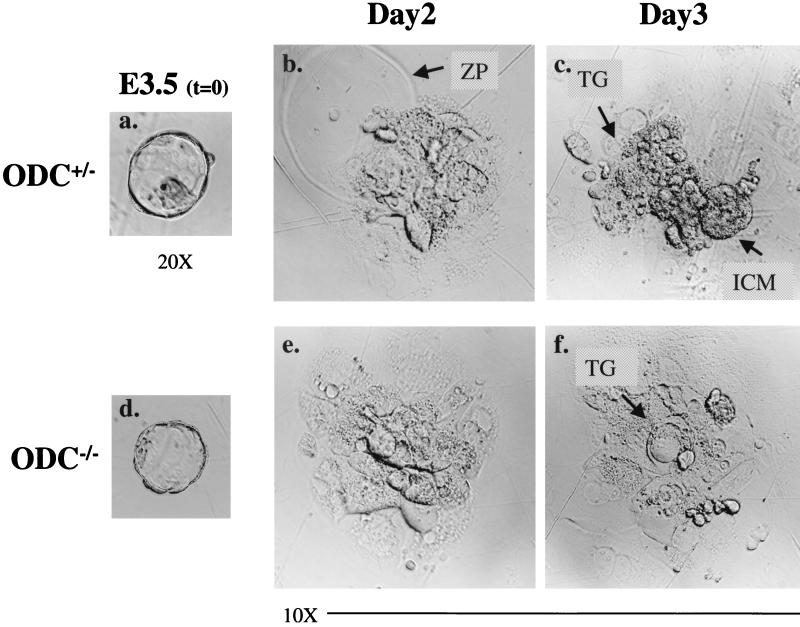
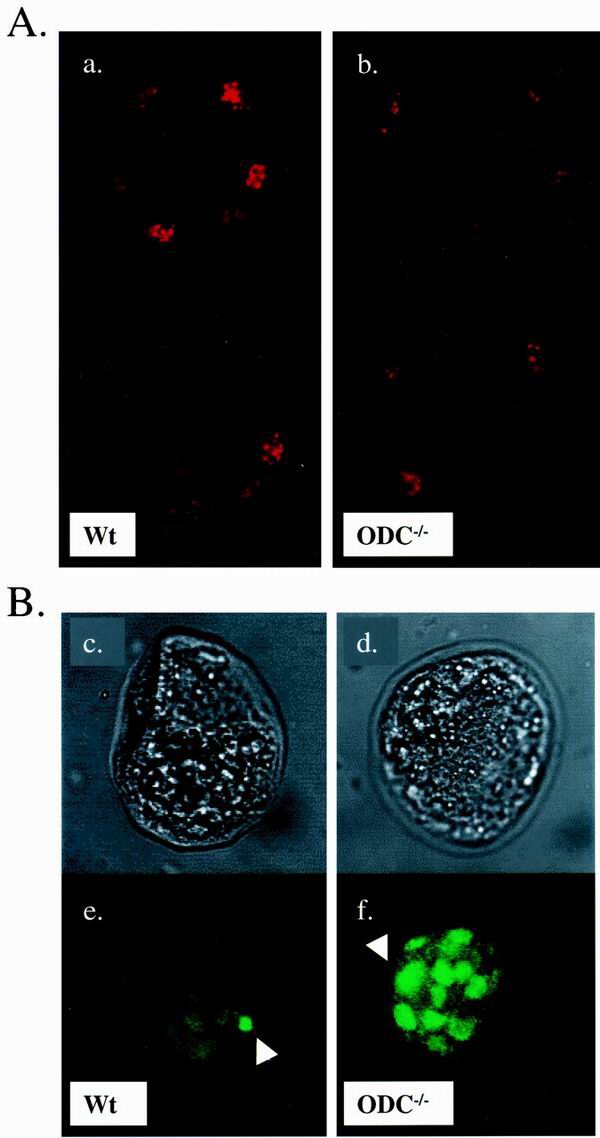
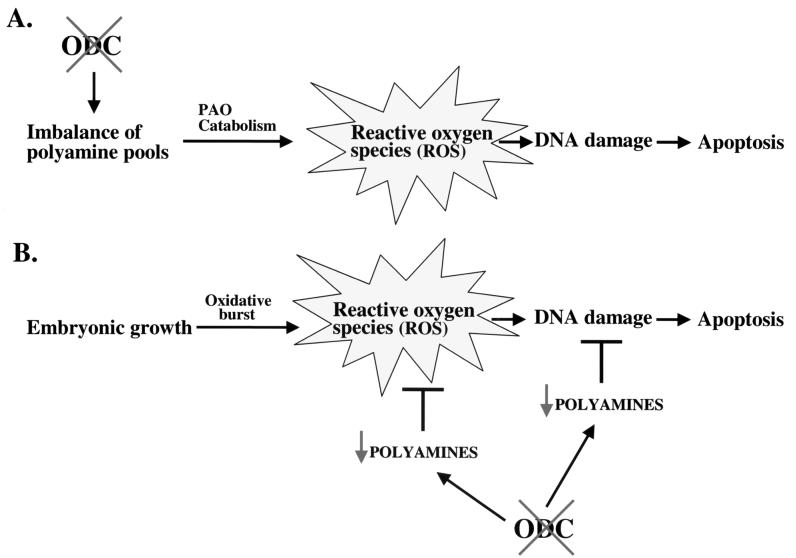
Overexpression and inhibitor studies have suggested that the c-Myc target gene for ornithine decarboxylase (ODC), the enzyme which converts ornithine to putrescine, plays an important role in diverse biological processes, including cell growth, differentiation, transformation, and apoptosis. To explore the physiological function of ODC in mammalian development, we generated mice harboring a disrupted ODC gene. ODC-heterozygous mice were viable, normal, and fertile. Although zygotic ODC is expressed throughout the embryo prior to implantation, loss of ODC did not block normal development to the blastocyst stage. Embryonic day E3.5 ODC-deficient embryos were capable of uterine implantation and induced maternal decidualization yet failed to develop substantially thereafter. Surprisingly, analysis of ODC-deficient blastocysts suggests that loss of ODC does not affect cell growth per se but rather is required for survival of the pluripotent cells of the inner cell mass. Therefore, ODC plays an essential role in murine development, and proper homeostasis of polyamine pools appears to be required for cell survival prior to gastrulation.
Figures
References
-
- Ajiro K, Yoda K, Utsumi K, Nishikawa Y. Alteration of cell cycle-dependent histone phosphorylations by okadaic acid. Induction of mitosis-specific H3 phosphorylation and chromatin condensation in mammalian interphase cells. J Biol Chem. 1996;271:13197–13201. - PubMed
-
- Askew D S, Ashmun R A, Simmons B C, Cleveland J L. Constitutive c-myc expression in an IL-3-dependent myeloid cell line suppresses cell cycle arrest and accelerates apoptosis. Oncogene. 1991;6:1915–1922. - PubMed
-
- Auvinen M, Paasinen A, Andersson L C, Holtta E. Ornithine decarboxylase activity is critical for cell transformation. Nature. 1992;360:355–358. - PubMed
-
- Balasundaram D, Tyagi A K. Polyamine-DNA nexus: structural ramifications and biological implications. Mol Cell Biochem. 1991;100:129–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials