

Checkpoint inhibition of the APC/C in HeLa cells is mediated by a complex of BUBR1, BUB3, CDC20, and MAD2
- PMID: 11535616
- PMCID: PMC2196190
- DOI: 10.1083/jcb.200102093
Checkpoint inhibition of the APC/C in HeLa cells is mediated by a complex of BUBR1, BUB3, CDC20, and MAD2
Abstract
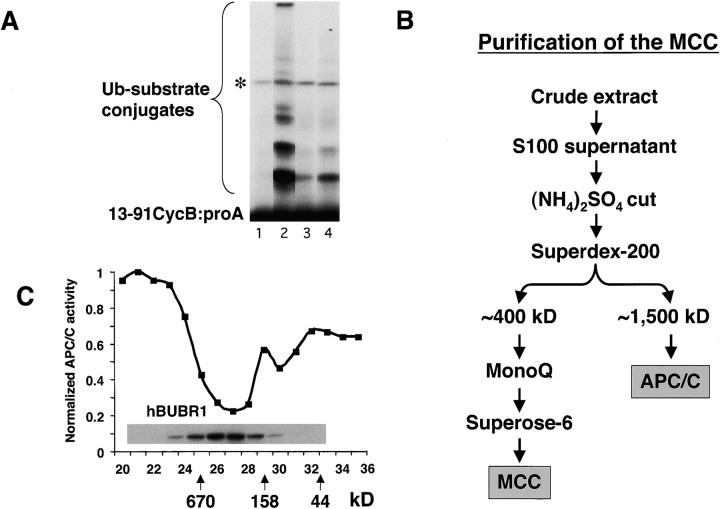
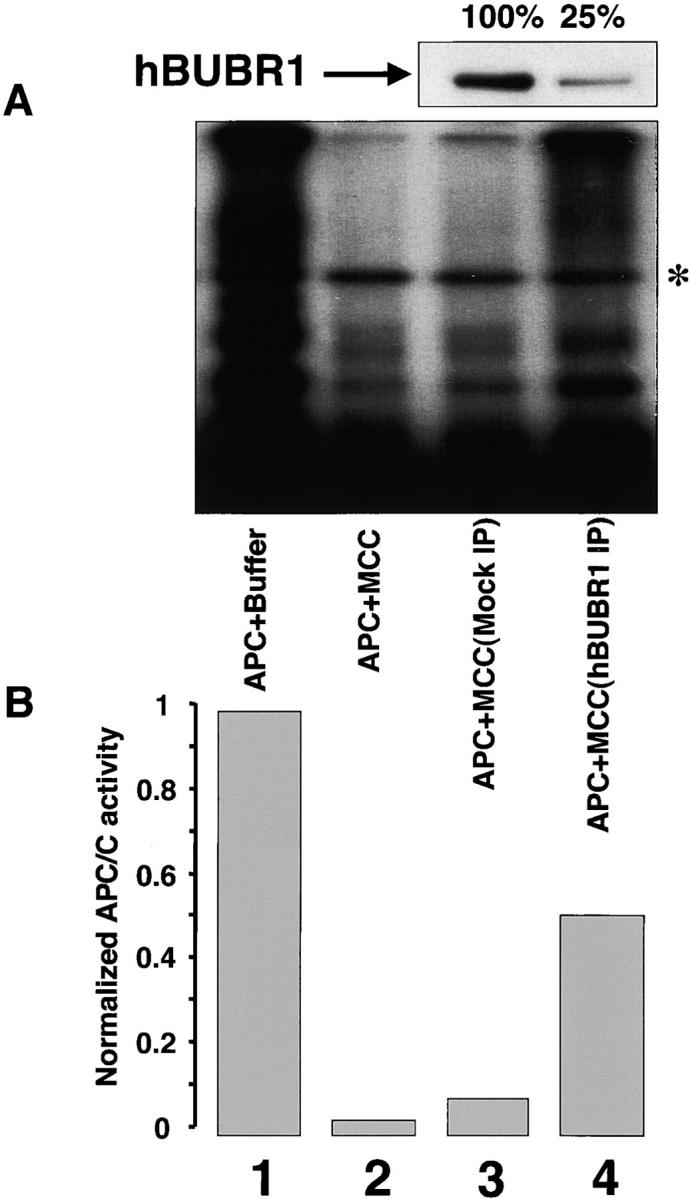
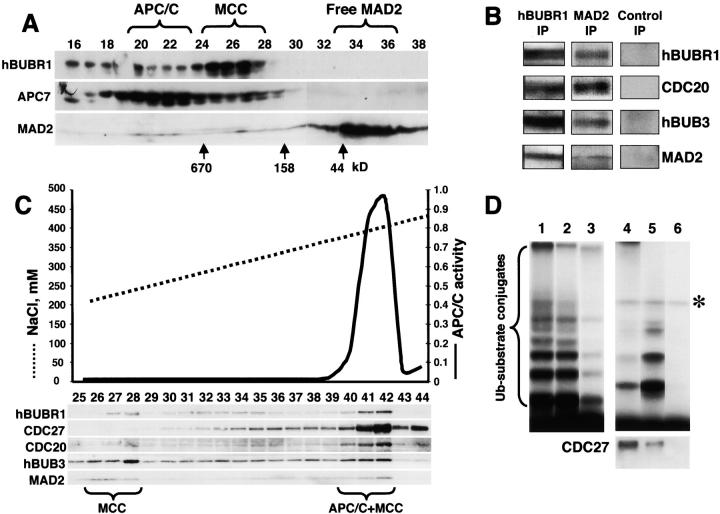
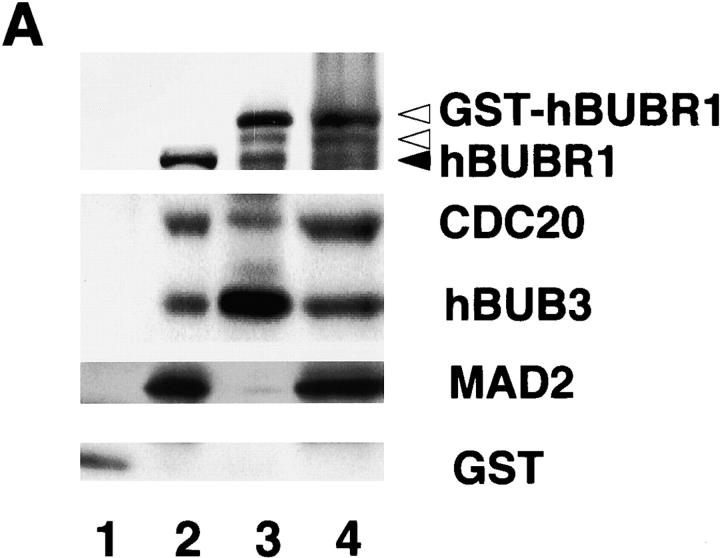
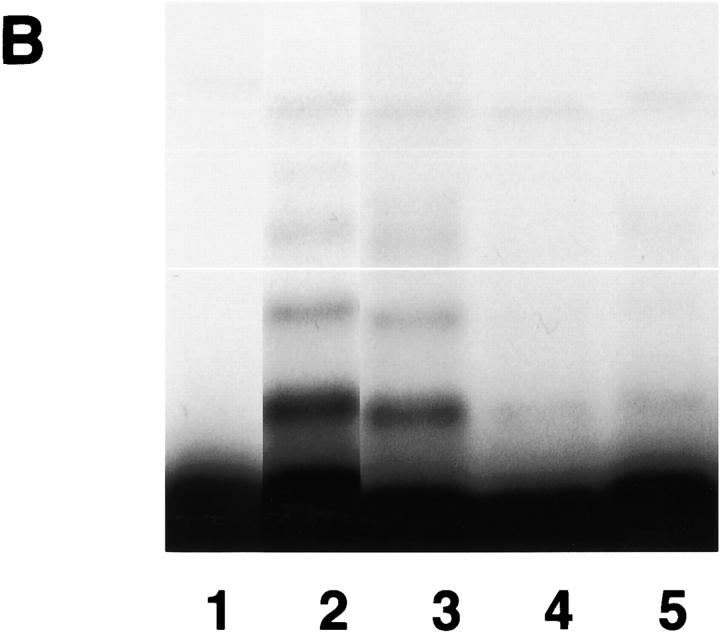
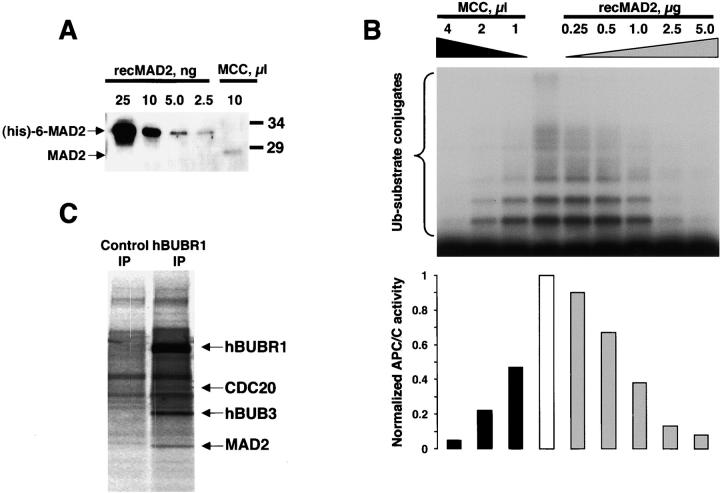
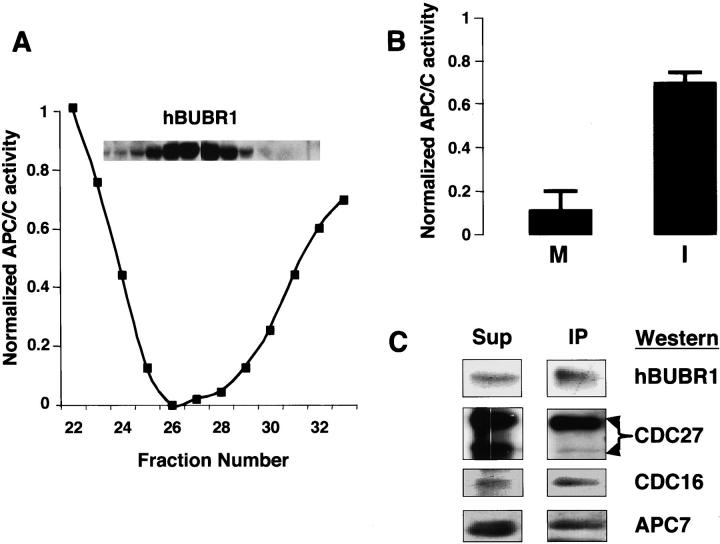
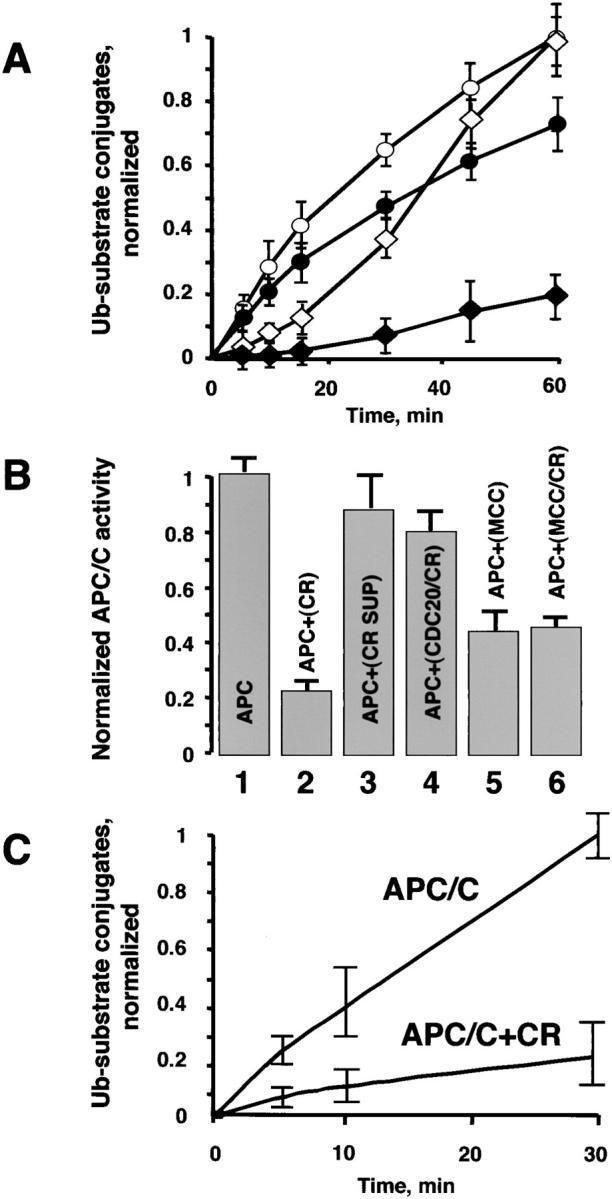
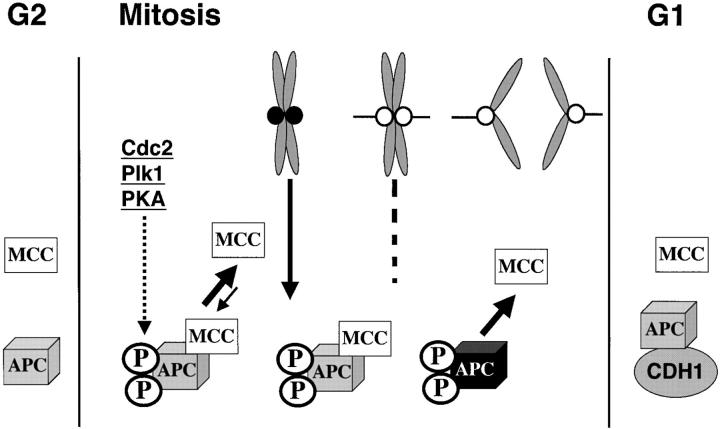
The mitotic checkpoint prevents cells with unaligned chromosomes from prematurely exiting mitosis by inhibiting the anaphase-promoting complex/cyclosome (APC/C) from targeting key proteins for ubiquitin-mediated proteolysis. We have examined the mechanism by which the checkpoint inhibits the APC/C by purifying an APC/C inhibitory factor from HeLa cells. We call this factor the mitotic checkpoint complex (MCC) as it consists of hBUBR1, hBUB3, CDC20, and MAD2 checkpoint proteins in near equal stoichiometry. MCC inhibitory activity is 3,000-fold greater than that of recombinant MAD2, which has also been shown to inhibit APC/C in vitro. Surprisingly, MCC is not generated from kinetochores, as it is also present and active in interphase cells. However, only APC/C isolated from mitotic cells was sensitive to inhibition by MCC. We found that the majority of the APC/C in mitotic lysates is associated with the MCC, and this likely contributes to the lag in ubiquitin ligase activity. Importantly, chromosomes can suppress the reactivation of APC/C. Chromosomes did not affect the inhibitory activity of MCC or the stimulatory activity of CDC20. We propose that the preformed interphase pool of MCC allows for rapid inhibition of APC/C when cells enter mitosis. Unattached kinetochores then target the APC/C for sustained inhibition by the MCC.
Figures
Comment in
-
A new view of the spindle checkpoint.J Cell Biol. 2001 Sep 3;154(5):909-11. doi: 10.1083/jcb.200108010. J Cell Biol. 2001. PMID: 11535614 Free PMC article. Review.
References
-
- Abrieu, A., J.A. Kahana, K.W. Wood, and D.W. Cleveland. 2000. CENP-E as an essential component of the mitotic checkpoint in vitro. Cell. 102:817–826. - PubMed
-
- Campbell, M.S., G.K.T. Chan, and T.J. Yen. 2001. Mitotic checkpoint proteins HsMAD1 and HsMAD2 are associated with nuclear pore complexes in interphase. J. Cell Sci. 114:953–963. - PubMed
-
- Charles, J.F., S.L. Jaspersen, R.L. Tinker-Kulberg, L. Hwang, A. Szidon, and D.O. Morgan. 1998. The Polo-related kinase Cdc5 activates and is destroyed by the mitotic cyclin destruction machinery in S. cerevisiae. Curr. Biol. 8:497–507. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
