

Searching sequence space for protein catalysts
- PMID: 11535813
- PMCID: PMC58511
- DOI: 10.1073/pnas.191159298
Searching sequence space for protein catalysts
Abstract
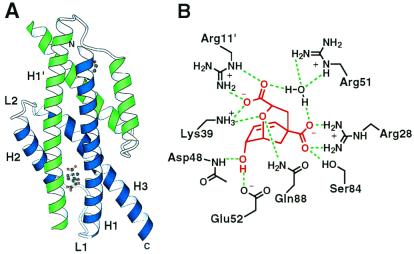
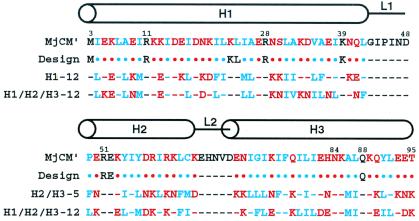
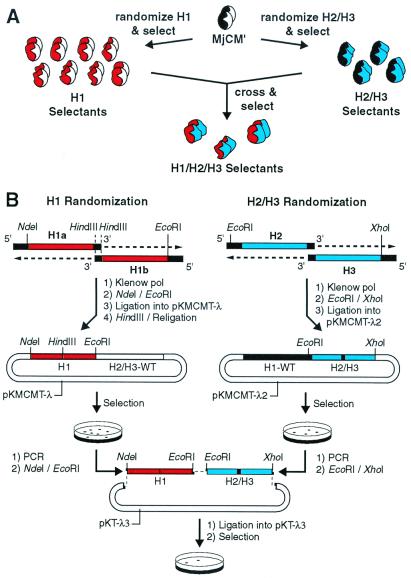
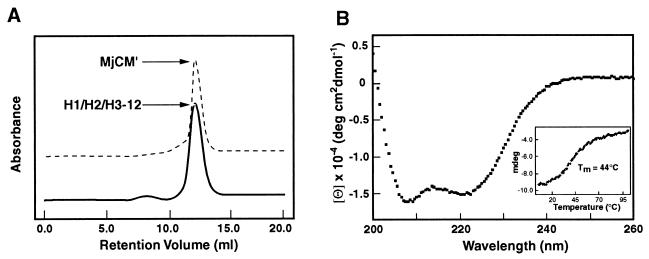
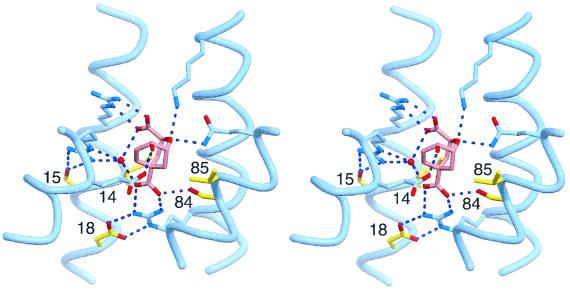
Genetic selection was used to explore the probability of finding enzymes in protein sequence space. Large degenerate libraries were prepared by replacing all secondary structure units in a dimeric, helical bundle chorismate mutase with simple binary-patterned modules based on a limited set of four polar and four nonpolar residues. Two-stage in vivo selection yielded catalytically active variants possessing biophysical and kinetic properties typical of the natural enzyme even though approximately 80% of the protein originates from the simplified modules and >90% of the protein consists of only eight different amino acids. This study provides a quantitative assessment of the number of sequences compatible with a given fold and implicates previously unidentified residues needed to form a functional active site. Given the extremely low incidence of enzymes in completely unbiased libraries, strategies that combine chemical information with genetic selection, like the one used here, may be generally useful in designing novel protein scaffolds with tailored activities.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
