

Human SMG-1, a novel phosphatidylinositol 3-kinase-related protein kinase, associates with components of the mRNA surveillance complex and is involved in the regulation of nonsense-mediated mRNA decay
- PMID: 11544179
- PMCID: PMC312771
- DOI: 10.1101/gad.913001
Human SMG-1, a novel phosphatidylinositol 3-kinase-related protein kinase, associates with components of the mRNA surveillance complex and is involved in the regulation of nonsense-mediated mRNA decay
Abstract
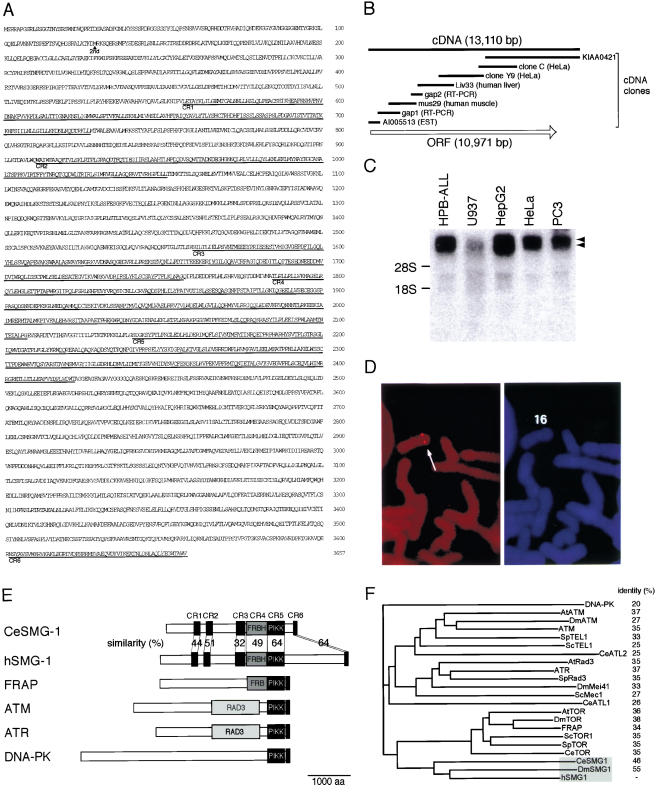
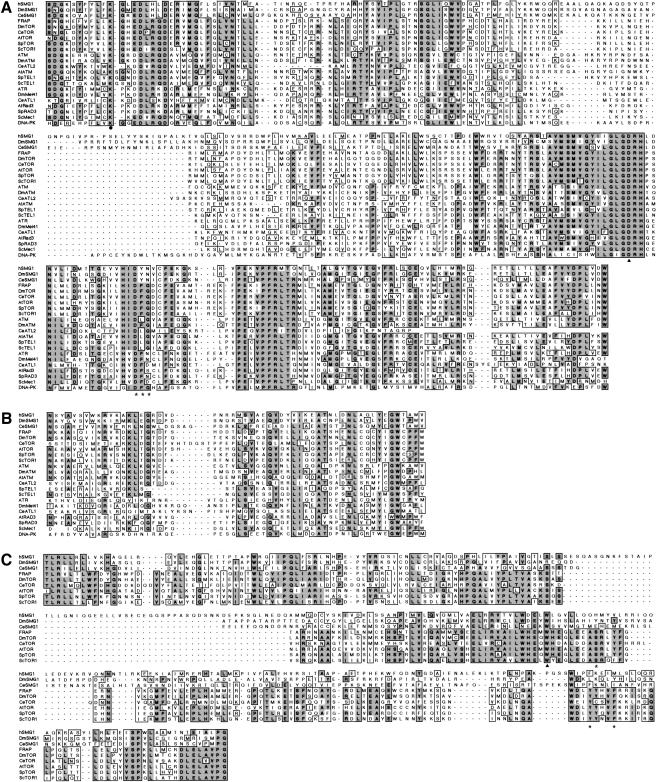
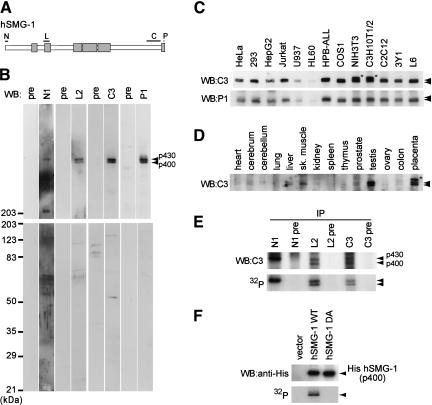
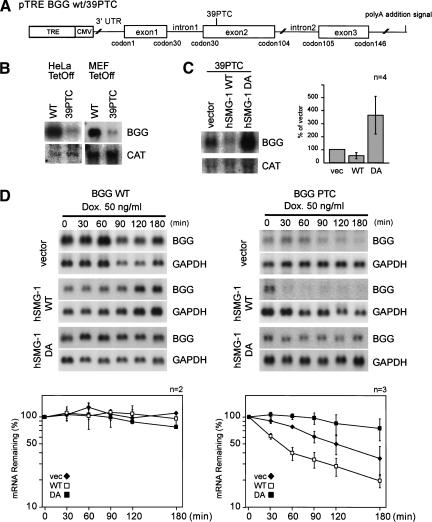
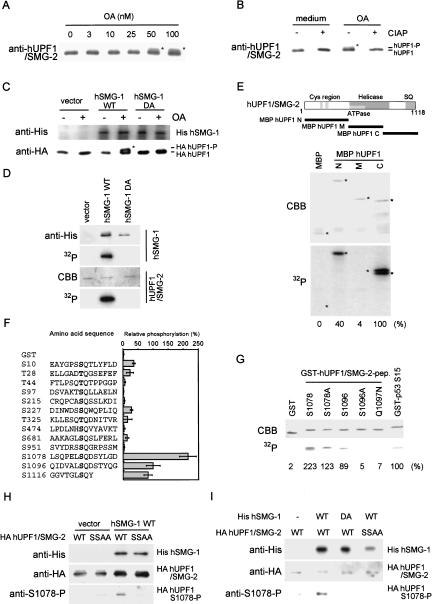
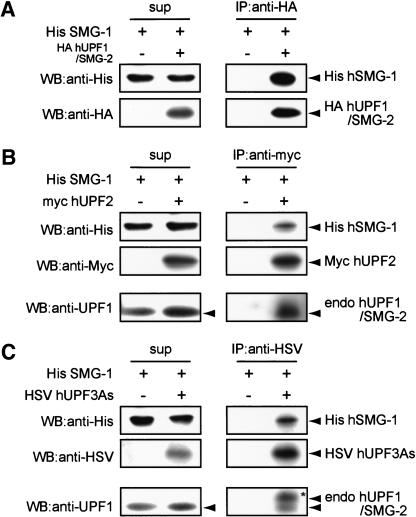
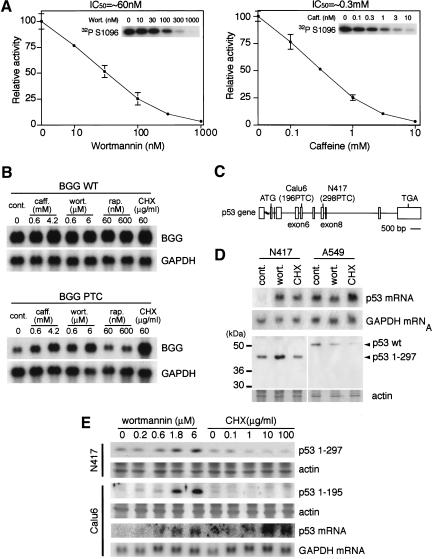
Nonsense-mediated mRNA decay (NMD) is a conserved surveillance mechanism that eliminates imperfect mRNAs that contain premature translation termination codons (PTCs) and code for nonfunctional or potentially harmful polypeptides. We show that a novel phosphatidylinositol 3-kinase-related protein kinase, hSMG-1, is a human ortholog of a product of Caenorhabditis elegans smg-1, one of seven smg genes involved in NMD. hSMG-1 phosphorylates hUPF1/SMG-2 in vivo and in vitro at specific serine residues in SQ motifs. hSMG-1 can associate with hUPF1/SMG-2 and other components of the surveillance complex. In particular, overexpression of a kinase-deficient point mutant of hSMG-1, hSMG-1-DA, results in a marked suppression of the PTC-dependent beta-globin mRNA degradation; whereas that of wild-type hSMG-1 enhances it. We also show that inhibitors of hSMG-1 induce the accumulation of truncated p53 proteins in human cancer cell lines with p53 PTC mutation. Taken together, we conclude that hSMG-1 plays a critical role in NMD through the direct phosphorylation of hUPF1/SMG-2 in the evolutionally conserved mRNA surveillance complex.
Figures
References
-
- Beelman CA, Stevens A, Caponigro G, LaGrandeur TE, Hatfield L, Fortner DM, Parker R. An essential component of the decapping enzyme required for normal rates of mRNA turnover. Nature. 1996;382:642–646. - PubMed
-
- Bodner SM, Minna JD, Jensen SM, D'Amico D, Carbone D, Mitsudomi T, Fedorko J, Buchhagen DL, Nau MM, Gazdar AF, et al. Expression of mutant p53 proteins in lung cancer correlates with the class of p53 gene mutation. Oncogene. 1992;7:743–749. - PubMed
-
- Brown EJ, Beal PA, Keith CT, Chen J, Shin TB, Schreiber SL. Control of p70 S6 kinase by kinase activity of FRAP in vivo. Nature. 1995;377:441–446. - PubMed
-
- Cali BM, Anderson P. mRNA surveillance mitigates genetic dominance in Caenorhabditis elegans. Mol Gen Genet. 1998;260:176–184. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous