

PDE1 encodes a P-type ATPase involved in appressorium-mediated plant infection by the rice blast fungus Magnaporthe grisea
- PMID: 11549759
- PMCID: PMC139447
- DOI: 10.1105/tpc.010056
PDE1 encodes a P-type ATPase involved in appressorium-mediated plant infection by the rice blast fungus Magnaporthe grisea
Abstract
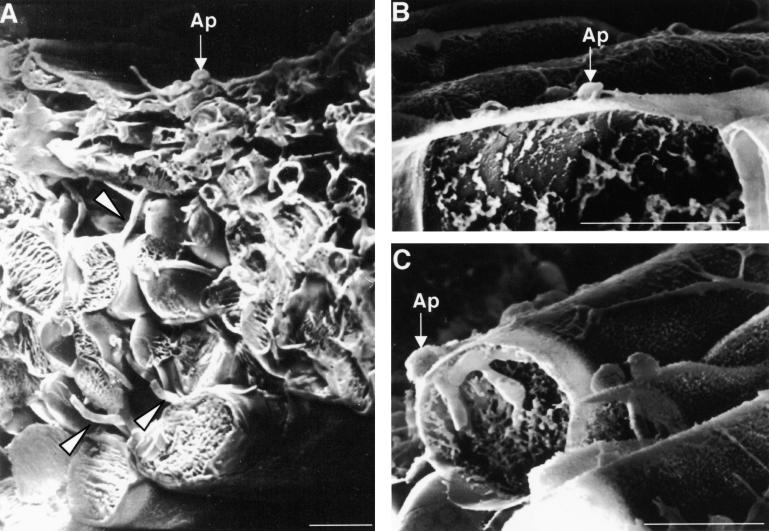
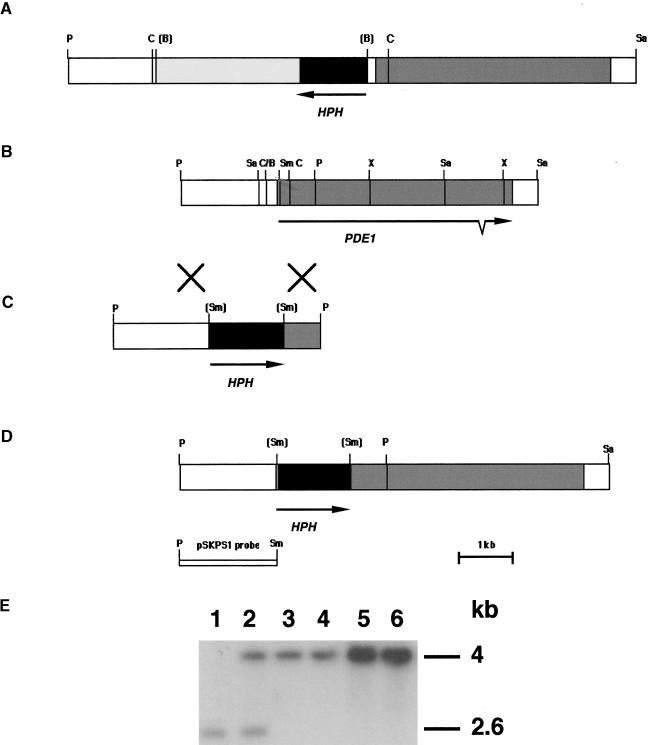
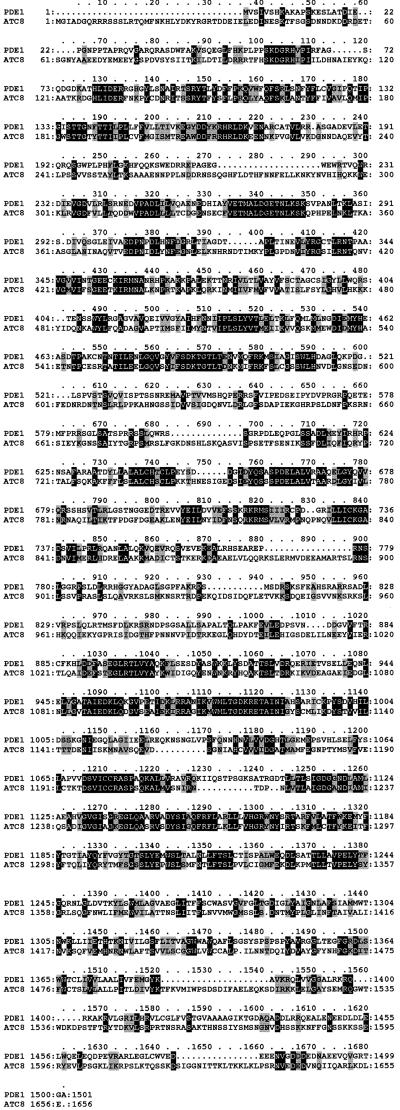
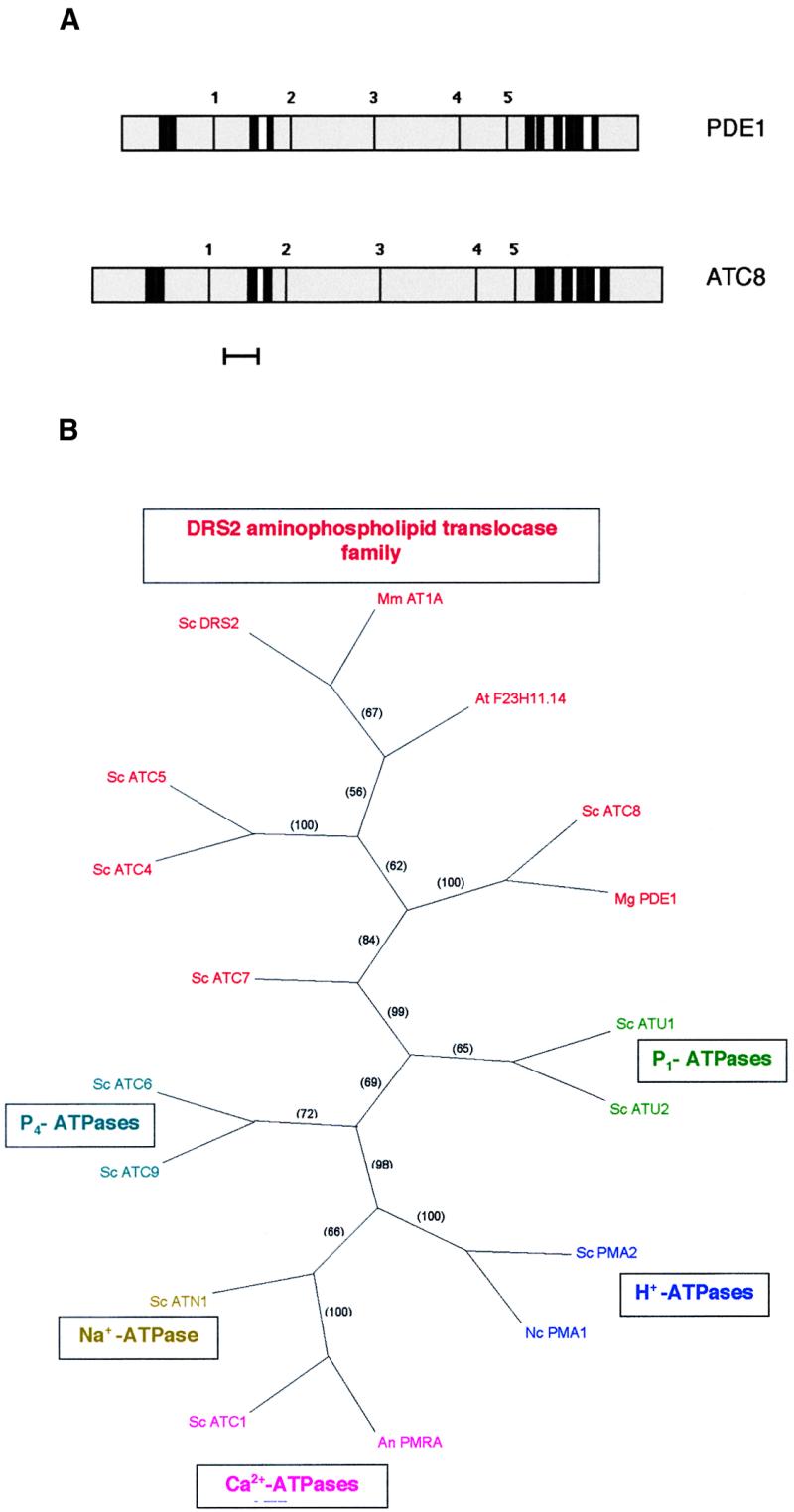
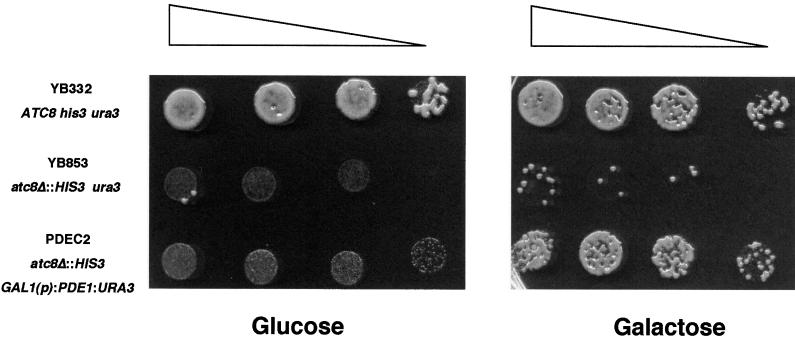
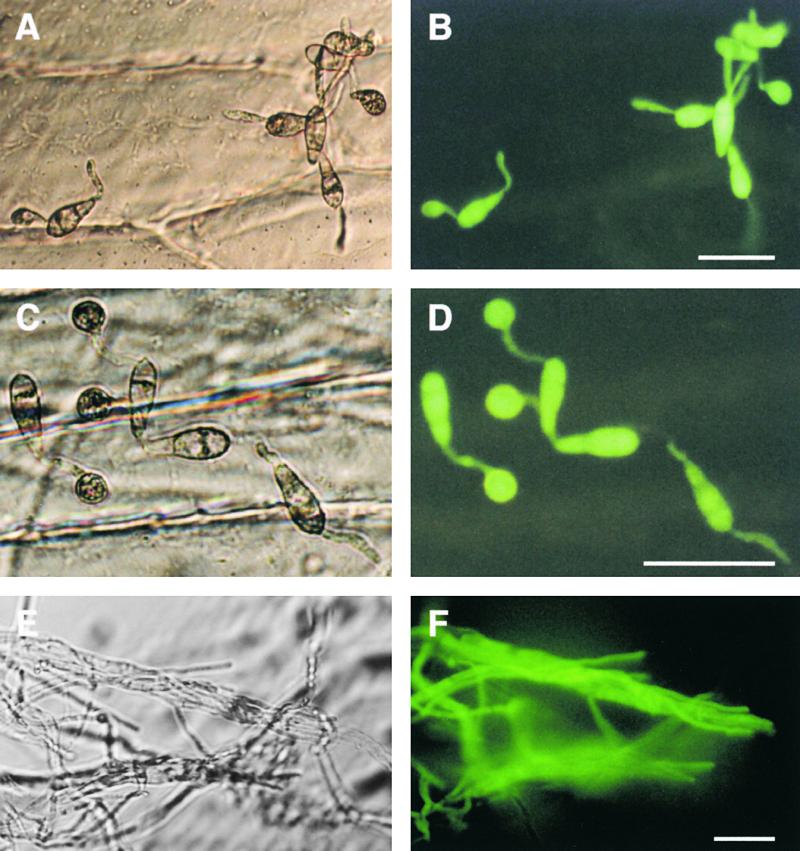
Plant infection by the rice blast fungus Magnaporthe grisea is brought about by the action of specialized infection cells called appressoria. These infection cells generate enormous turgor pressure, which is translated into an invasive force that allows a narrow penetration hypha to breach the plant cuticle. The Magnaporthe pde1 mutant was identified previously by restriction enzyme-mediated DNA integration mutagenesis and is impaired in its ability to elaborate penetration hyphae. Here we report that the pde1 mutation is the result of an insertion into the promoter of a P-type ATPase-encoding gene. Targeted gene disruption confirmed the role of PDE1 in penetration hypha development and pathogenicity but highlighted potential differences in PDE1 regulation in different Magnaporthe strains. The predicted PDE1 gene product was most similar to members of the aminophospholipid translocase group of P-type ATPases and was shown to be a functional homolog of the yeast ATPase gene ATC8. Spatial expression studies showed that PDE1 is expressed in germinating conidia and developing appressoria. These findings implicate the action of aminophospholipid translocases in the development of penetration hyphae and the proliferation of the fungus beyond colonization of the first epidermal cell.
Figures


References
-
- Altschul, S.F., Gish, W., Miller, W., Myers, C.W., and Lipman, D.L. (1990). Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
-
- Ashrafi, K., Farazi, T.A., and Gordon, J.I. (1998). A role for Saccharomyces cerevisiae fatty acid activation protein 4 in regulating protein N-myristoylation during entry into stationary phase. J. Biol. Chem. 273 25864–25874. - PubMed
-
- Balhadère, P.V., Foster, A.J., and Talbot, N.J. (1999). Identification of pathogenicity mutants of the rice blast fungus Magnaporthe grisea by insertional mutagenesis. Mol. Plant-Microbe Interact. 12 129–142.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
