

A 90-kD phospholipase D from tobacco binds to microtubules and the plasma membrane
- PMID: 11549769
- PMCID: PMC139457
- DOI: 10.1105/tpc.010114
A 90-kD phospholipase D from tobacco binds to microtubules and the plasma membrane
Abstract
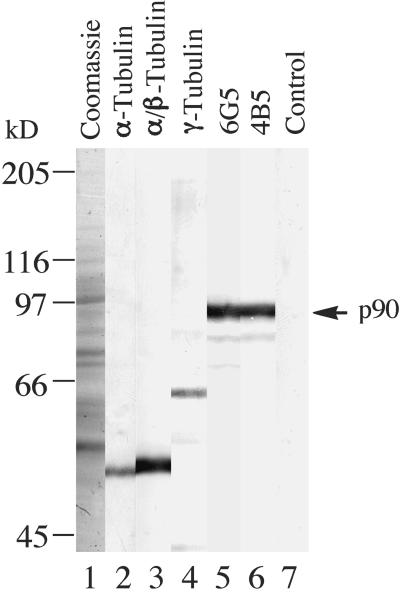
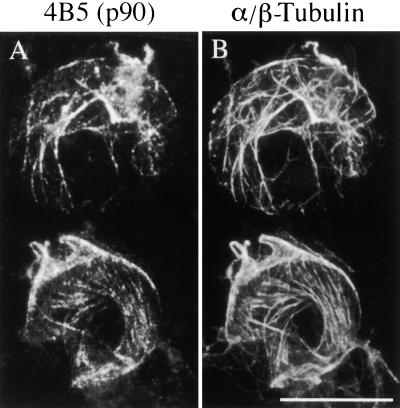
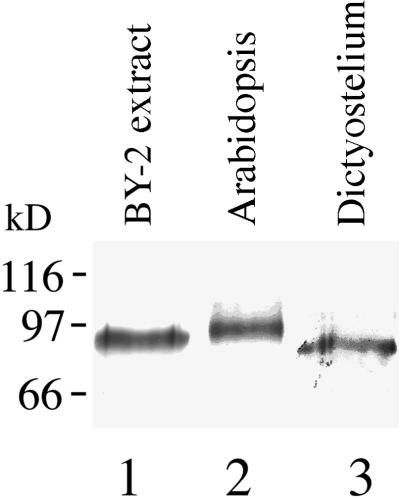
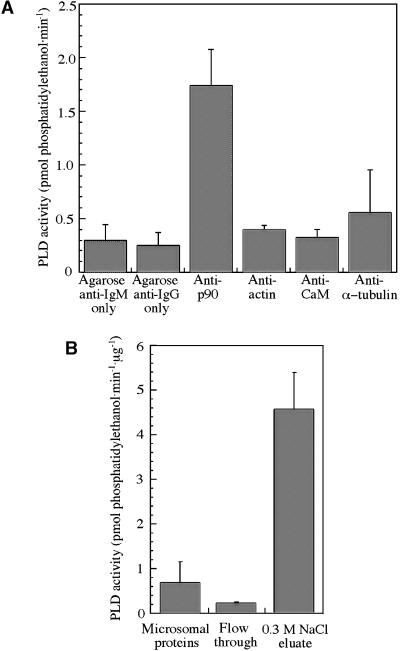
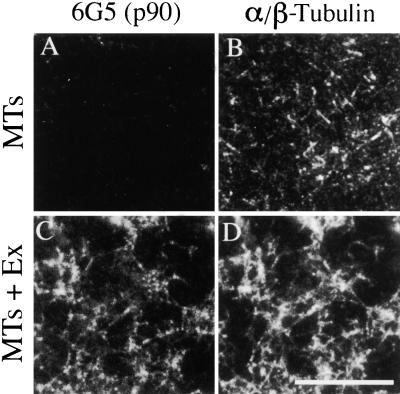
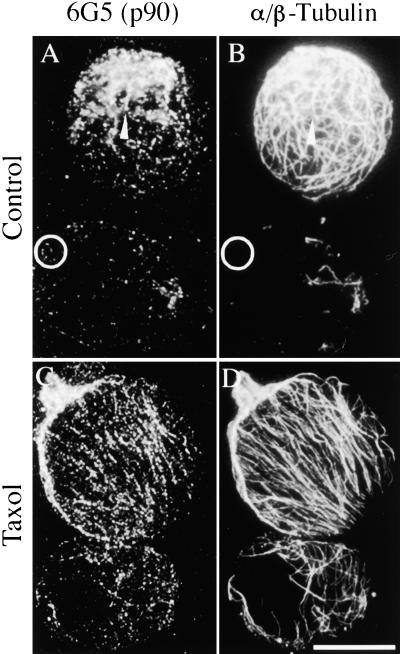
The organization of microtubule arrays in the plant cell cortex involves interactions with the plasma membrane, presumably through protein bridges. We have used immunochemistry and monoclonal antibody 6G5 against a candidate bridge protein, a 90-kD tubulin binding protein (p90) from tobacco BY-2 membranes, to characterize the protein and isolate the corresponding gene. Screening an Arabidopsis cDNA expression library with the antibody 6G5 produced a partial clone encoding phospholipase D (PLD), and a full-length gene was obtained by sequencing a corresponding expressed sequence tag clone. The predicted protein of 857 amino acids contains the active sites of a phospholipid-metabolizing enzyme and a Ca(2+)-dependent lipid binding domain and is identical to Arabidopsis PLD delta. Two amino acid sequences obtained by Edman degradation of the tobacco p90 are identical to corresponding segments of a PLD sequence from tobacco. Moreover, immunoprecipitation using the antibody 6G5 and tobacco BY-2 protein extracts gave significant PLD activity, and PLD activity of tobacco BY-2 membrane proteins was enriched 6.7-fold by tubulin-affinity chromatography. In a cosedimentation assay, p90 bound and decorated microtubules. In immunofluorescence microscopy of intact tobacco BY-2 cells or lysed protoplasts, p90 colocalized with cortical microtubules, and taxol-induced microtubule bundling was accompanied by corresponding reorganization of p90. Labeling of p90 remained along the plasma membrane when microtubules were depolymerized, although detergent extraction abolished the labeling. Therefore, p90 is a specialized PLD that associates with membranes and microtubules, possibly conveying hormonal and environmental signals to the microtubule cytoskeleton.
Figures



References
-
- Akashi, T., and Shibaoka, H. (1991). Involvement of transmembrane proteins in the association of cortical microtubules with the plasma membrane in tobacco BY-2 cells. J. Cell Sci. 98 169–174.
-
- Akashi, T., Kawasaki, S., and Shibaoka, H. (1990). Stabilization of cortical microtubules by the cell wall in cultured tobacco cells: Effects of extensin on the cold-stability of cortical microtubules. Planta 182 363–369. - PubMed
-
- Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
-
- Asada, T., and Collings, D. (1997). Molecular motors in higher plants. Trends Plant Sci. 2 29–37.
-
- Assmann, S.M., and Baskin, T.I. (1998). The function of guard cells does not require an intact array of cortical microtubules. J. Exp. Bot. 49 163–170.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
