

Mannheimia haemolytica leukotoxin activates a nonreceptor tyrosine kinase signaling cascade in bovine leukocytes, which induces biological effects
- PMID: 11553552
- PMCID: PMC98743
- DOI: 10.1128/IAI.69.10.6131-6139.2001
Mannheimia haemolytica leukotoxin activates a nonreceptor tyrosine kinase signaling cascade in bovine leukocytes, which induces biological effects
Abstract
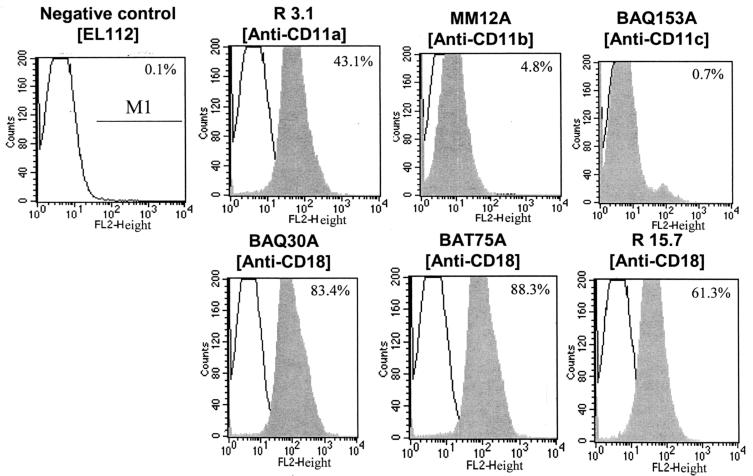
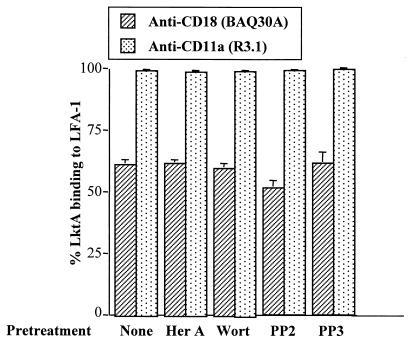
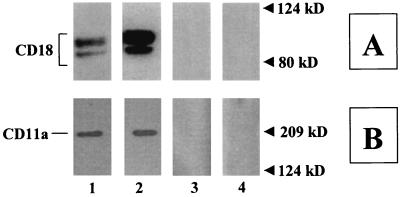
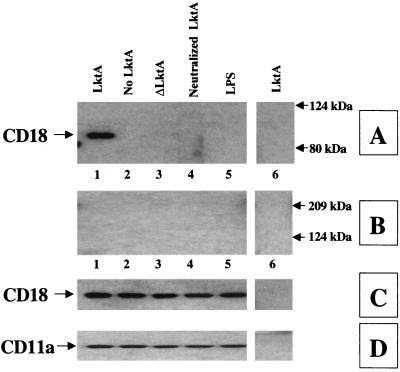
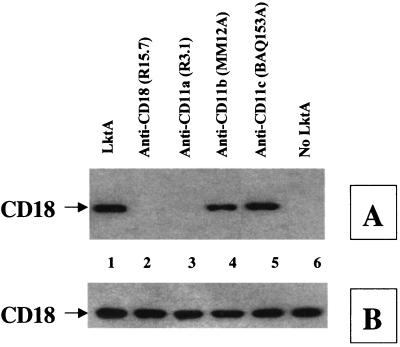
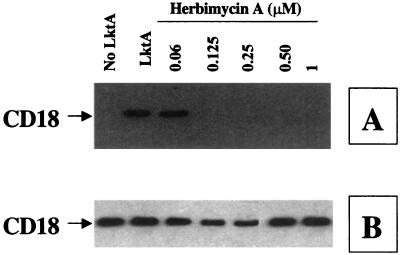
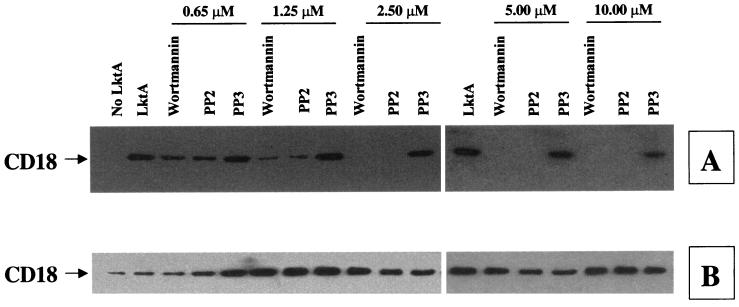
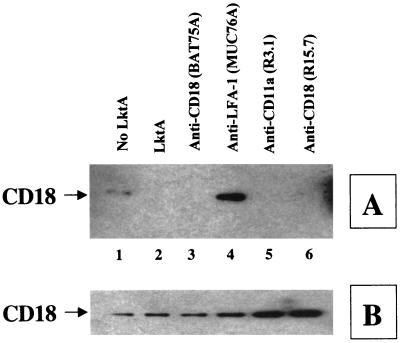
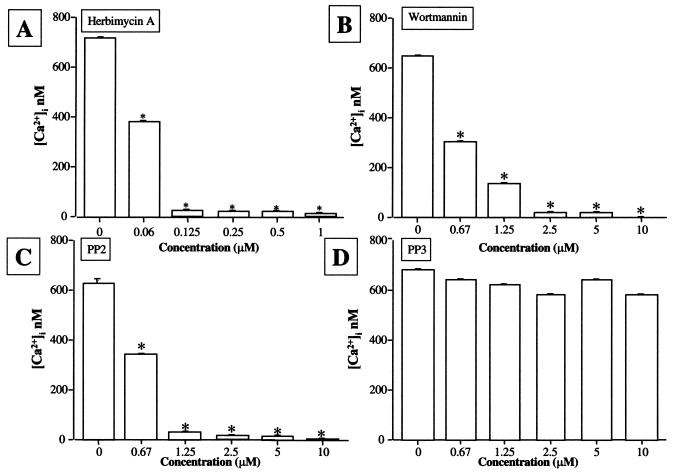
The leukotoxin (LktA) produced by Mannheimia haemolytica binds to bovine lymphocyte function-associated antigen 1 (LFA-1) and induces biological effects in bovine leukocytes in a cellular and species-specific fashion. We have previously shown that LktA also binds to porcine LFA-1 without eliciting any effects. These findings suggest that the specificity of LktA effects must entail both binding to LFA-1 and activation of signaling pathways which are present in bovine leukocytes. However, the signaling pathways leading to biological effects upon LktA binding to LFA-1 have not been characterized. In this context, several reports have indicated that ligand binding to LFA-1 results in activation of a nonreceptor tyrosine kinase (NRTK) signaling cascade. We designed experiments with the following objectives: (i) to determine whether LktA binding to LFA-1 leads to activation of NRTKs, (ii) to examine whether LktA-induced NRTK activation is target cell specific, and (iii) to determine whether LktA-induced NRTK activation is required for biological effects. We used a biologically inactive mutant leukotoxin (DeltaLktA) for comparison with LktA. Our results indicate that LktA induces tyrosine phosphorylation (TP) of the CD18 tail of LFA-1 in bovine leukocytes. The DeltaLktA mutant does not induce TP of the CD18 tail, albeit binding to bovine LFA-1. LktA-induced TP of the CD18 tail was attenuated by an NRTK inhibitor, herbimycin A; a phosphatidylinositol 3'-kinase (PI 3-kinase) inhibitor, wortmannin; and a Src kinase inhibitor, PP2, in a concentration-dependent manner. Furthermore, LktA induces TP of the CD18 tail in bovine, but not porcine, leukocytes. Moreover, LktA-induced intracellular calcium ([Ca2+]i) elevation was also inhibited by herbimycin A, wortmannin, and PP2. Thus, our data represent the first evidence that binding of LktA to bovine LFA-1 induces a species-specific NRTK signaling cascade involving PI 3-kinase and Src kinases and that this signaling cascade is required for LktA-induced biological effects.
Figures
References
-
- Albert R K, Embree L J, McFeely J E, Hickstein D D. Expression and function of β2 integrins on alveolar macrophages from human and nonhuman primates. Am J Respir Cell Mol Biol. 1992;7:182–189. - PubMed
-
- Allan E M, Wiseman A, Gibbs H A, Selman I E. Pasteurella species isolated from the bovine respiratory tract and their antimicrobial sensitivity patterns. Vet Rec. 1985;117:629–631. - PubMed
-
- Ambagala T C, Ambagala A P, Srikumaran S. The leukotoxin of Pasteurella haemolytica binds to β2 integrins on bovine leukocytes. FEMS Microbiol Lett. 1999;179:161–167. - PubMed
-
- Beaty C D, Franklin T L, Uhera Y, Wilson C B. Lipopolysaccharide-induced cytokine production in human monocytes: role of tyrosine phosphorylation in transmembrane signal tranduction. Eur J Immunol. 1994;24:1278–1284. - PubMed
-
- Clemston K J. Introduction: integrins, dynamic cell receptors. Cell Mol Life Sci. 1998;54:499–501.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
