

Folding and subunit assembly of photoreceptor peripherin/rds is mediated by determinants within the extracellular/intradiskal EC2 domain: implications for heterogeneous molecular pathologies
- PMID: 11553636
- PMCID: PMC1360209
- DOI: 10.1074/jbc.M107511200
Folding and subunit assembly of photoreceptor peripherin/rds is mediated by determinants within the extracellular/intradiskal EC2 domain: implications for heterogeneous molecular pathologies
Abstract
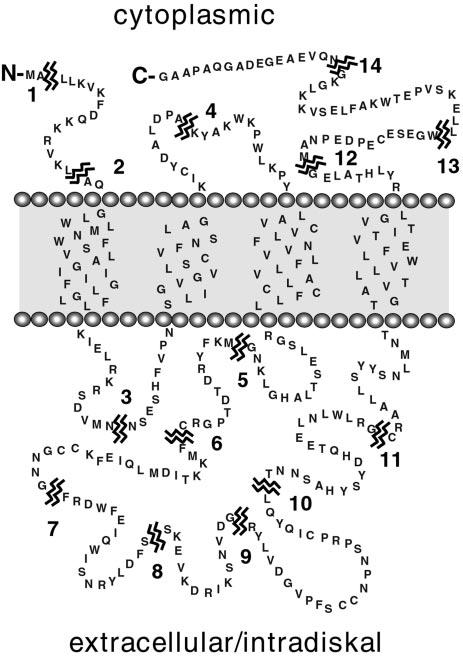
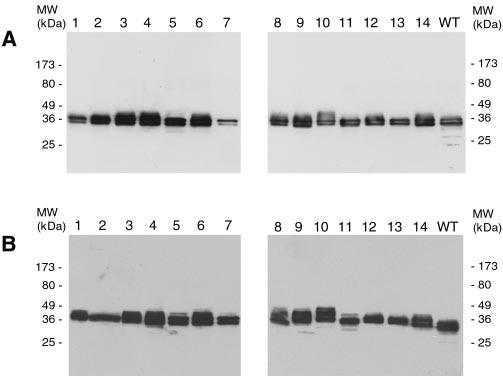
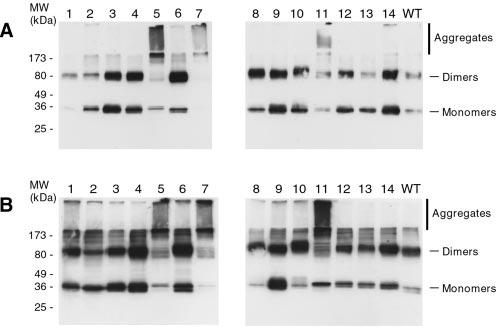
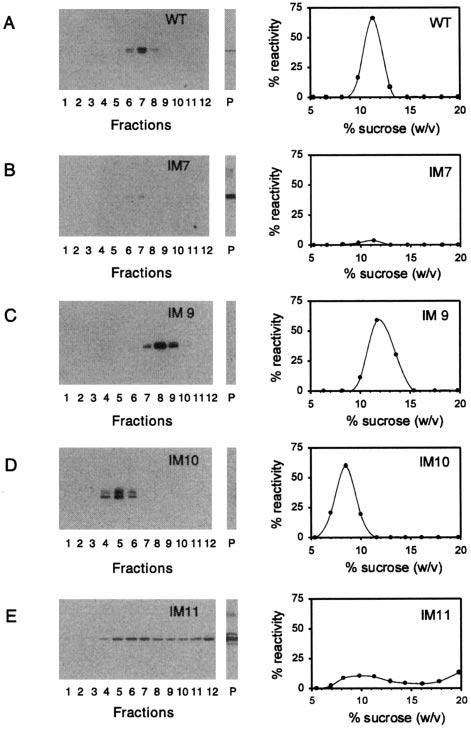
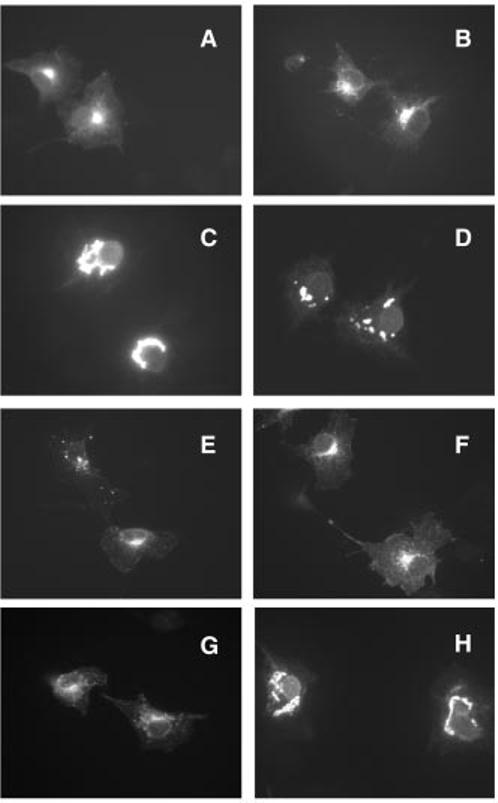
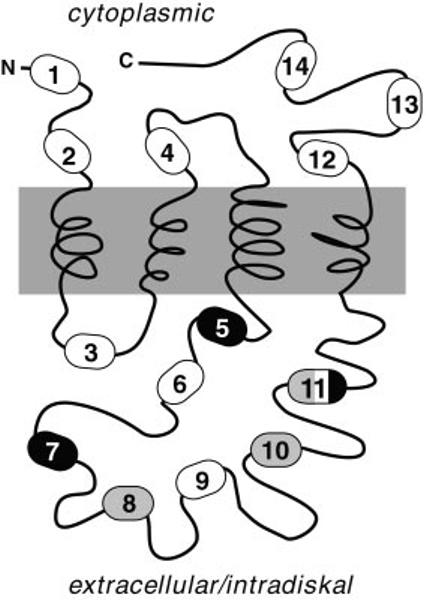
Peripherin/rds is an integral membrane protein required for the elaboration of rod and cone photoreceptor outer segments in the vertebrate retina; it causes a surprising variety of progressive retinal degenerations in humans and dysmorphic photoreceptors in murine models if defective or absent. (Peripherin/rds is also known as photoreceptor peripherin, peripherin/rds, rds/peripherin, rds, and peripherin-2.) Peripherin/rds appears to act as a structural element in outer segment architecture. However, neither its function at the molecular level nor its role in retinal disease processes are well understood. This report initiates a systematic investigation of protein domain structure and function by examining the molecular and cellular consequences of a series of 14 insertional mutations distributed throughout the polypeptide sequence. Protein expression, disulfide bonding, sedimentation velocity, and subcellular localization of the COS-1 cell-expressed mutant variants were examined to test the hypothesis that protein folding and tetrameric subunit assembly are mediated primarily by EC2, a conserved extracellular/intradiskal domain. Protein folding and tetrameric subunit assembly were not affected by insertion of either an uncharged dipeptide (GA) or a highly charged hendecapeptide (GDYKDDDDKAA) into any one of nine sites residing outside of EC2 as assayed by nonreducing Western blot analysis, sedimentation velocity, and subcellular localization. In contrast, insertions at five positions within the EC2 domain did cause either gross protein misfolding (two sites) or a reduction in protein sedimentation coefficient (two sites) or both (one site). These results indicate that although the vast majority of extramembranous polypeptide sequence makes no measurable contribution to protein folding and tetramerization, discrete regions within the EC2 domain do contain determinants for normal subunit assembly. These findings raise the possibility that multiple classes of structural perturbation are produced by inherited defects in peripherin/rds and contribute to the observed heterogeneity of retinal disease phenotypes.
Figures
Similar articles
-
Cysteine residues of photoreceptor peripherin/rds: role in subunit assembly and autosomal dominant retinitis pigmentosa.Biochemistry. 1998 Jan 13;37(2):680-5. doi: 10.1021/bi972036i. Biochemistry. 1998. PMID: 9425091
-
Uncoupling of photoreceptor peripherin/rds fusogenic activity from biosynthesis, subunit assembly, and targeting: a potential mechanism for pathogenic effects.J Biol Chem. 2004 Sep 17;279(38):39958-67. doi: 10.1074/jbc.M403943200. Epub 2004 Jul 13. J Biol Chem. 2004. PMID: 15252042 Free PMC article.
-
Disulfide-mediated oligomerization of Peripherin/Rds and Rom-1 in photoreceptor disk membranes. Implications for photoreceptor outer segment morphogenesis and degeneration.J Biol Chem. 2000 Feb 25;275(8):5370-8. doi: 10.1074/jbc.275.8.5370. J Biol Chem. 2000. PMID: 10681511
-
Role of peripherin/rds in vertebrate photoreceptor architecture and inherited retinal degenerations.Int Rev Cytol. 2006;253:131-75. doi: 10.1016/S0074-7696(06)53004-9. Int Rev Cytol. 2006. PMID: 17098056 Review.
-
Role of subunit assembly in autosomal dominant retinitis pigmentosa linked to mutations in peripherin 2.Novartis Found Symp. 2004;255:95-112; discussion 113-6, 177-8. Novartis Found Symp. 2004. PMID: 14750599 Review.
Cited by
-
Molecular basis for photoreceptor outer segment architecture.Prog Retin Eye Res. 2016 Nov;55:52-81. doi: 10.1016/j.preteyeres.2016.05.003. Epub 2016 Jun 1. Prog Retin Eye Res. 2016. PMID: 27260426 Free PMC article. Review.
-
Multistep peripherin-2/rds self-assembly drives membrane curvature for outer segment disk architecture and photoreceptor viability.Proc Natl Acad Sci U S A. 2020 Feb 25;117(8):4400-4410. doi: 10.1073/pnas.1912513117. Epub 2020 Feb 10. Proc Natl Acad Sci U S A. 2020. PMID: 32041874 Free PMC article.
-
Insights into the mechanisms of macular degeneration associated with the R172W mutation in RDS.Hum Mol Genet. 2014 Jun 15;23(12):3102-14. doi: 10.1093/hmg/ddu014. Epub 2014 Jan 25. Hum Mol Genet. 2014. PMID: 24463884 Free PMC article.
-
The B3 Subunit of the Cone Cyclic Nucleotide-gated Channel Regulates the Light Responses of Cones and Contributes to the Channel Structural Flexibility.J Biol Chem. 2016 Apr 15;291(16):8721-34. doi: 10.1074/jbc.M115.696138. Epub 2016 Feb 18. J Biol Chem. 2016. PMID: 26893377 Free PMC article.
-
Photoreceptor renewal: a role for peripherin/rds.Int Rev Cytol. 2002;217:183-225. doi: 10.1016/s0074-7696(02)17015-x. Int Rev Cytol. 2002. PMID: 12019563 Free PMC article. Review.
References
-
- Young RW. Invest. Ophthalmol. Vis. Sci. 1976;15:700–725. - PubMed
-
- Molday RS, Hicks D, Molday L. Invest. Ophthalmol. Vis. Sci. 1987;28:50–61. - PubMed
-
- Travis GH, Brennan MB, Danielson PE, Kozak CA, Sutcliffe JG. Nature. 1989;338:70–73. - PubMed
-
- Sanyal S, Jansen HG. Neurosci. Lett. 1981;21:23–26. - PubMed
-
- Kohl S, Giddings I, Besch D, Apfelstedt-Sylla E, Zrenner E, Wissinger B. Acta Anat. (Basel) 1998;162:75–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
