

Involvement of integrin alpha(v)beta(3) and cell adhesion molecule L1 in transendothelial migration of melanoma cells
- PMID: 11553709
- PMCID: PMC59705
- DOI: 10.1091/mbc.12.9.2699
Involvement of integrin alpha(v)beta(3) and cell adhesion molecule L1 in transendothelial migration of melanoma cells
Abstract
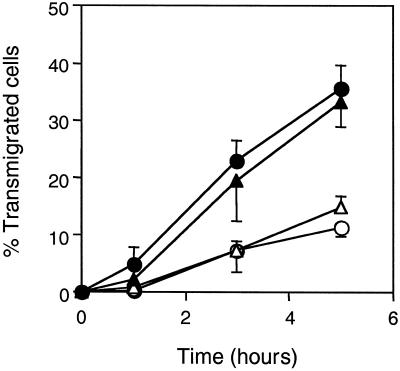
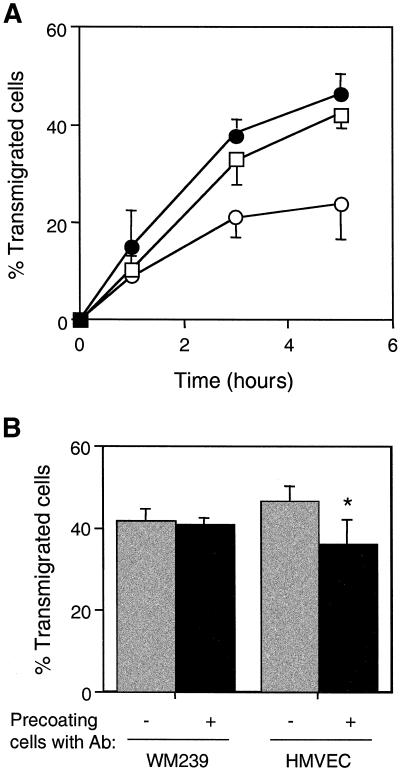
Tumor metastasis involves many stage-specific adhesive interactions. The expression of several cell adhesion molecules, notably the integrin alpha(v)beta(3), has been associated with the metastatic potential of tumor cells. In this study, we used a novel in vitro assay to examine the role of alpha(v)beta(3) in the transmigration of melanoma cells through a monolayer of human lung microvascular endothelial cells. Confocal microscopy revealed the presence of the integrin alpha(v)beta(3) on melanoma membrane protrusions and pseudopods penetrating the endothelial junction. alpha(v)beta(3) was also enriched in heterotypic contacts between endothelial cells and melanoma cells. Transendothelial migration of melanoma cells was inhibited by either a cyclic Arg-Gly-Asp peptide or the anti-alpha(v)beta(3) monoclonal antibody LM609. Although both platelet endothelial cell adhesion molecule-1 and L1 are known to bind integrin alpha(v)beta(3), only L1 serves as a potential ligand for alpha(v)beta(3) during melanoma transendothelial migration. Also, polyclonal antibodies against L1 partially inhibited the transendothelial migration of melanoma cells. However, addition of both L1 and alpha(v)beta(3) antibodies did not show additive effects, suggesting that they are components of the same adhesion system. Together, the data suggest that interactions between the integrin alpha(v)beta(3) on melanoma cells and L1 on endothelial cells play an important role in the transendothelial migration of melanoma cells.
Figures






Similar articles
-
Platelet-endothelial cell adhesion molecule-1 (CD31) redistributes from the endothelial junction and is not required for the transendothelial migration of melanoma cells.Clin Exp Metastasis. 2000;18(6):527-32. doi: 10.1023/a:1011884807746. Clin Exp Metastasis. 2000. PMID: 11592310
-
Human neural cell adhesion molecule L1 and rat homologue NILE are ligands for integrin alpha v beta 3.J Cell Biol. 1996 Feb;132(3):475-85. doi: 10.1083/jcb.132.3.475. J Cell Biol. 1996. PMID: 8636223 Free PMC article.
-
The L1 adhesion molecule supports alpha v beta 3-mediated migration of human tumor cells and activated T lymphocytes.Biochem Biophys Res Commun. 1997 Mar 6;232(1):236-9. doi: 10.1006/bbrc.1997.6265. Biochem Biophys Res Commun. 1997. PMID: 9125139
-
Cell-cell interactions during transendothelial migration of tumor cells.Microsc Res Tech. 1998 Nov 1;43(3):265-75. doi: 10.1002/(SICI)1097-0029(19981101)43:3<265::AID-JEMT9>3.0.CO;2-Z. Microsc Res Tech. 1998. PMID: 9840805 Review.
-
Cellular adhesion pathways and metastatic potential of human melanoma.Cancer Biol Ther. 2002 Sep-Oct;1(5):459-65. doi: 10.4161/cbt.1.5.158. Cancer Biol Ther. 2002. PMID: 12496470 Review.
Cited by
-
L1CAM: a major driver for tumor cell invasion and motility.Cell Adh Migr. 2012 Jul-Aug;6(4):374-84. doi: 10.4161/cam.20832. Epub 2012 Jul 1. Cell Adh Migr. 2012. PMID: 22796939 Free PMC article. Review.
-
Platelet-endothelial cell adhesion molecule-1 (CD31) redistributes from the endothelial junction and is not required for the transendothelial migration of melanoma cells.Clin Exp Metastasis. 2000;18(6):527-32. doi: 10.1023/a:1011884807746. Clin Exp Metastasis. 2000. PMID: 11592310
-
Connexin 43 mediated gap junctional communication enhances breast tumor cell diapedesis in culture.Breast Cancer Res. 2005;7(4):R522-34. doi: 10.1186/bcr1042. Epub 2005 May 13. Breast Cancer Res. 2005. PMID: 15987459 Free PMC article.
-
L1CAM malfunction in the nervous system and human carcinomas.Cell Mol Life Sci. 2010 Jul;67(14):2425-37. doi: 10.1007/s00018-010-0339-1. Epub 2010 Mar 17. Cell Mol Life Sci. 2010. PMID: 20237819 Free PMC article. Review.
-
Role of the endothelium during tumor cell metastasis: is the endothelium a barrier or a promoter for cell invasion and metastasis?J Biophys. 2008;2008:183516. doi: 10.1155/2008/183516. Epub 2009 Mar 5. J Biophys. 2008. PMID: 20107573 Free PMC article.
References
-
- Albelda SM, Mette SA, Elder DE, Stewart R, Damjanovich L, Herlyn M, Buck CA. Integrin distribution in malignant melanoma: association of the β3 subunit with tumor progression. Cancer Res. 1990;50:6757–6764. - PubMed
-
- Ashman LK, Aylett GW, Cambareri AC, Cole SR. Different epitopes of the CD31 antigen identified by monoclonal antibodies: cell type-specific patterns of expression. Tissue Antigens. 1991;38:199–207. - PubMed
-
- Bani MR, Rak J, Adachi D, Wiltshire R, Trent JM, Kerbel RS, Ben-David Y. Multiple features of advanced melanoma recapitulated in tumorigenic variants of early stage (radial growth phase) human melanoma cell lines: evidence for a dominant phenotype. Cancer Res. 1996;56:3075–3086. - PubMed
-
- Basbaum C, Werb Z. Focalized proteolysis: spatial and temporal regulation of extracellular matrix degradation at the cell surface. Curr Opin Cell Biol. 1996;8:731–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
