

SUMO-1 modification required for transformation by adenovirus type 5 early region 1B 55-kDa oncoprotein
- PMID: 11553772
- PMCID: PMC58726
- DOI: 10.1073/pnas.191361798
SUMO-1 modification required for transformation by adenovirus type 5 early region 1B 55-kDa oncoprotein
Abstract
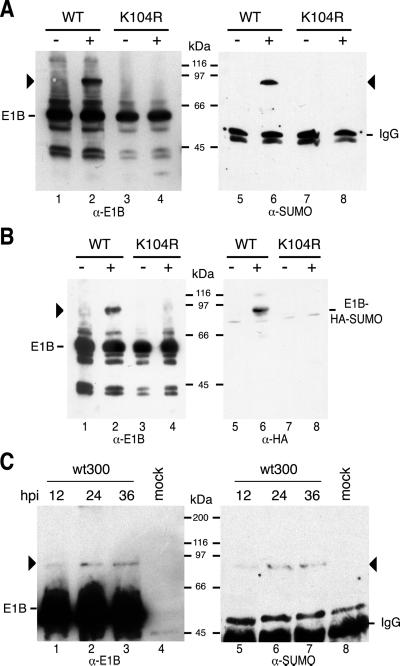
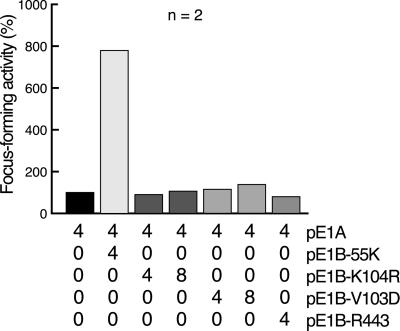
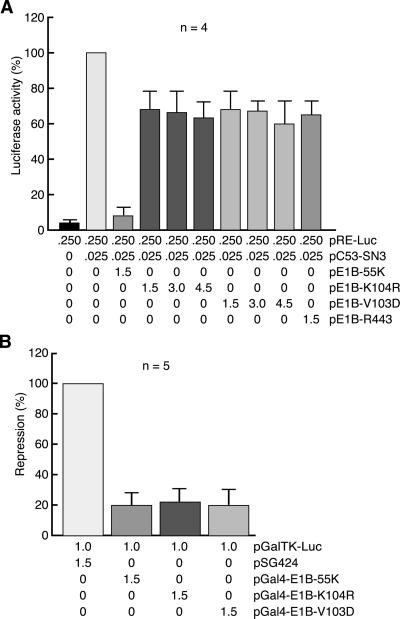
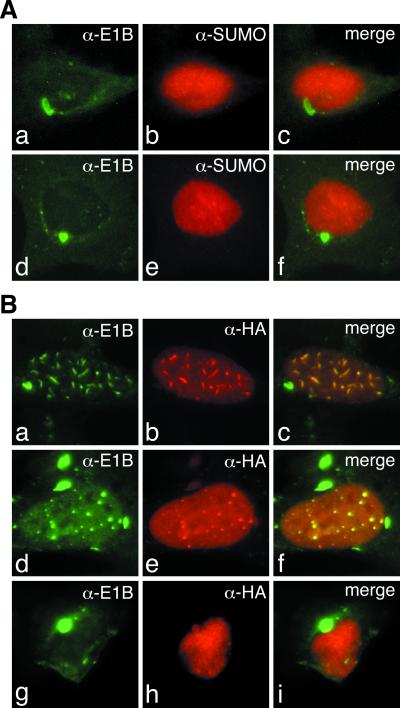
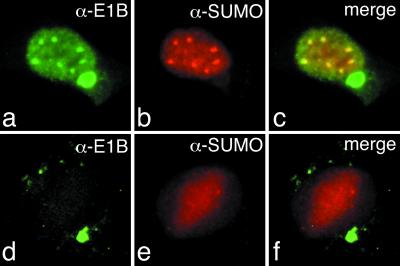
SUMO-1 is a small ubiquitin-related modifier protein that is covalently linked to many cellular and viral protein targets. Modification by SUMO-1 is proposed to play a role in protein targeting and/or stability. We show here that adenovirus type 5 early region 1B 55-kDa (E1B-55kDa) oncoprotein can be covalently modified by SUMO-1 in vivo through a major attachment site comprising a single lysine residue at amino acid position 104. The sequence surrounding this lysine matches the proposed PsiKxE consensus motif required for SUMO-1 conjugation. A single mutation (K104R) that abolishes SUMOylation of E1B-55kDa dramatically reduces the ability of the adenovirus type 5 protein to transform primary baby rat kidney cells in cooperation with E1A and to inhibit p53-mediated transactivation. Overexpression of SUMO-1 in adenovirus type 5 E1A/E1B-55kDa-transformed baby rat kidney cells causes the relocalization of E1B-55kDa from the cytoplasm to the nucleus, where it accumulates with SUMO-1 in dot- or track-like structures. Significantly, when SUMO-1 is ectopically expressed in transformed rat cells no effect on the cytoplasmic localization of the E1B-K104R mutant protein is observed. Our results demonstrate that SUMO-1 modification is required for transformation by adenovirus type 5 E1B-55kDa and provide further evidence for the idea that this posttranslational modification plays a role in protein targeting to specific subcellular sites.
Figures

References
-
- Shenk T. In: Virology. Fields B N, Knipe D M, Howley P M, editors. Vol. 2. New York: Lippincott-Raven; 1996. pp. 2111–2148.
-
- Barker D D, Berk A J. Virology. 1987;156:107–121. - PubMed
-
- Sarnow P, Ho Y S, Williams J, Levine A J. Cell. 1982;28:387–394. - PubMed
-
- Yew P R, Berk A J. Nature (London) 1992;357:82–85. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
