

The S4-S5 linker couples voltage sensing and activation of pacemaker channels
- PMID: 11553787
- PMCID: PMC58720
- DOI: 10.1073/pnas.201250598
The S4-S5 linker couples voltage sensing and activation of pacemaker channels
Abstract
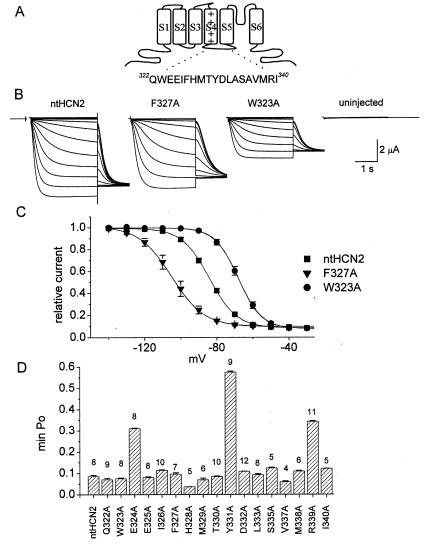
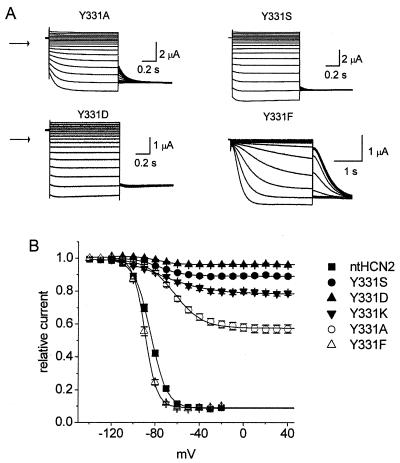
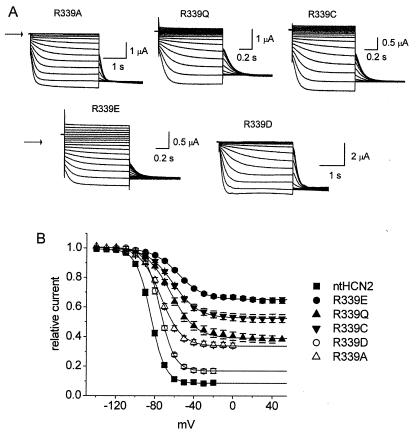
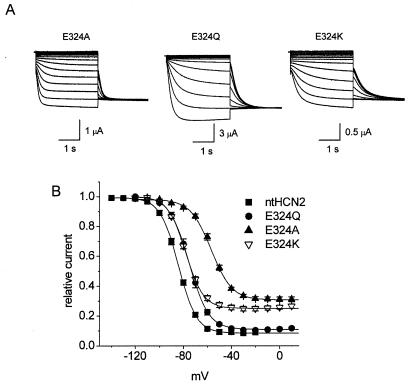
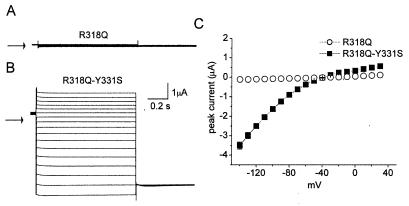
Voltage-gated channels are normally opened by depolarization and closed by repolarization of the membrane. Despite sharing significant sequence homology with voltage-gated K(+) channels, the gating of hyperpolarization-activated, cyclic-nucleotide-gated (HCN) pacemaker channels has the opposite dependence on membrane potential: hyperpolarization opens, whereas depolarization closes, these channels. The mechanism and structural basis of the process that couples voltage sensor movement to HCN channel opening and closing is not understood. On the basis of our previous studies of a mutant HERG (human ether-a-go-go-related gene) channel, we hypothesized that the intracellular linker that connects the fourth and fifth transmembrane domains (S4-S5 linker) of HCN channels might be important for channel gating. Here, we used alanine-scanning mutagenesis of the HCN2 S4-S5 linker to identify three residues, E324, Y331, and R339, that when mutated disrupted normal channel closing. Mutation of a basic residue in the S4 domain (R318Q) prevented channel opening, presumably by disrupting S4 movement. However, channels with R318Q and Y331S mutations were constitutively open, suggesting that these channels can open without a functioning S4 domain. We conclude that the S4-S5 linker mediates coupling between voltage sensing and HCN channel activation. Our findings also suggest that opening of HCN and related channels corresponds to activation of a gate located near the inner pore, rather than recovery of channels from a C-type inactivated state.
Figures
References
-
- DiFrancesco D, Noma A, Trautwein W. Pflügers Arch. 1979;381:271–279. - PubMed
-
- DiFrancesco D. Annu Rev Physiol. 1993;55:455–472. - PubMed
-
- Pape H-C. Annu Rev Physiol. 1996;58:299–327. - PubMed
-
- Ludwig A, Zong X, Jeglitsch M, Hofmann F, Biel M. Nature (London) 1998;393:587–591. - PubMed
-
- Santoro B, Liu D T, Yao H, Bartsch D, Kandel E R, Siegelbaum S A, Tibbs G R. Cell. 1998;93:717–729. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
