

Review
doi: 10.1093/nar/29.18.3757.
Homing endonucleases: structural and functional insight into the catalysts of intron/intein mobility
Affiliations
- PMID: 11557808
- PMCID: PMC55915
- DOI: 10.1093/nar/29.18.3757
Item in Clipboard
Review
Homing endonucleases: structural and functional insight into the catalysts of intron/intein mobility
Nucleic Acids Res.
.
Abstract
Homing endonucleases confer mobility to their host intervening sequence, either an intron or intein, by catalyzing a highly specific double-strand break in a cognate allele lacking the intervening sequence. These proteins are characterized by their ability to bind long DNA target sites (14-40 bp) and their tolerance of minor sequence changes in these sites. A wealth of biochemical and structural data has been generated for these enzymes over the past few years. Herein we review our current understanding of homing endonucleases, including their diversity and evolution, DNA-binding and catalytic mechanisms, and attempts to engineer them to bind novel DNA substrates.
Figures
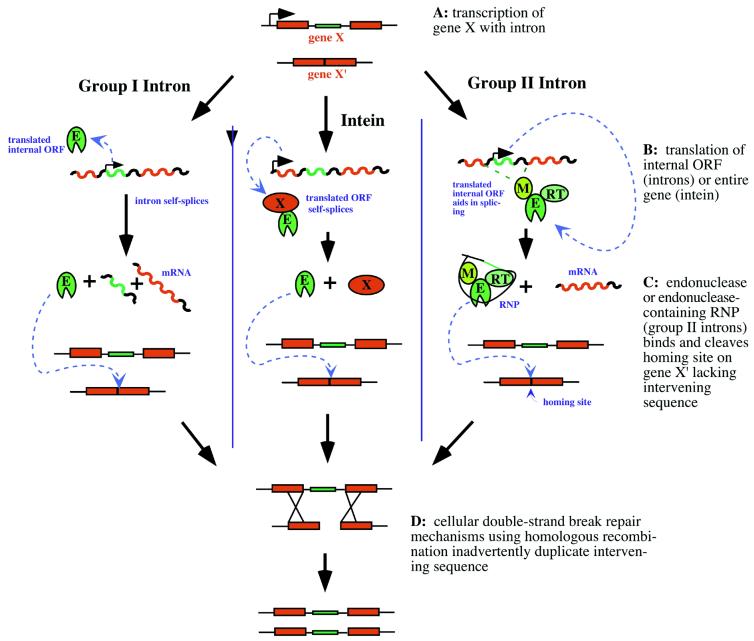
Homing mechanisms of group
I introns (left), inteins (center) and group II introns (right)
in which the intervening sequence of gene X is duplicated in its
cognate allele, gene X′. Mobile ORFs
and encoded products are green; host gene exons and products are
red; other nucleic acid sequence is black. E, endonuclease; M, maturase;
RT, reverse-transcriptase; RNP, ribonucleoprotein.
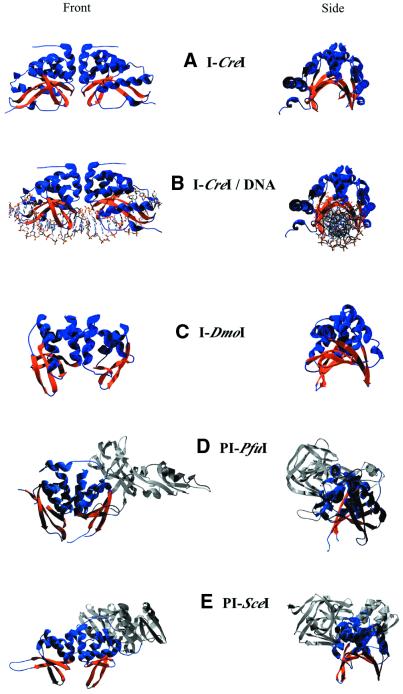
Structures of LAGLIDADG family
members. Endonuclease domains are blue with orange β-sheets
showing DNA-binding saddle. Other intein domains are gray. (A)
I-CreI homodimer. (B) I-CreI
with DNA. (C) I-DmoI monomer.
(D) PI-PfuI monomer. (E)
PI-SceI monomer.
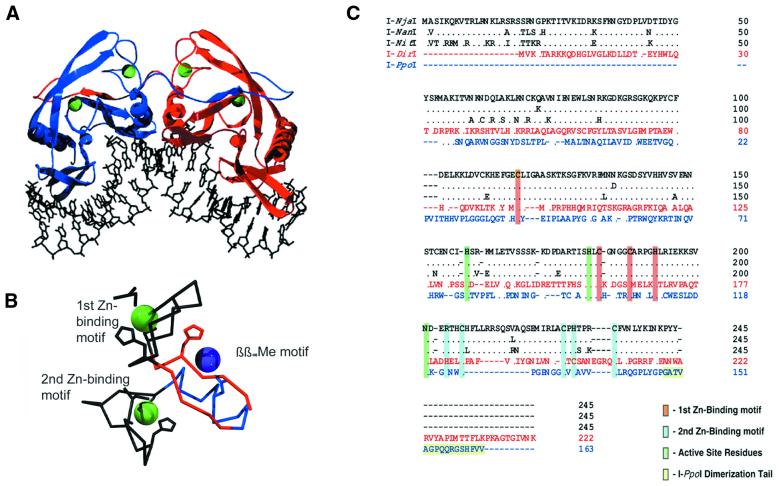
His-Cys box family. (A)
I-PpoI homodimer bound to its DNA target site.
Note ‘domain-swapped’ C-terminal tails that form
much of the dimer interface. Zinc atoms are green. (B)
Core metal-binding motifs in I-PpoI. Zinc ions
are green; magnesium ion is purple. β-strands
and α-helix of the conserved ββαMe
motif are red and blue, respectively. (C) Primary
sequence alignment of known His-Cys box family members including
I-NjaI, I-NanI, I-NitI,
I-DirI and I-PpoI. The C-terminal
dimerization tail of I-PpoI and the highly conserved
zinc-binding and active sight residues are highlighted.
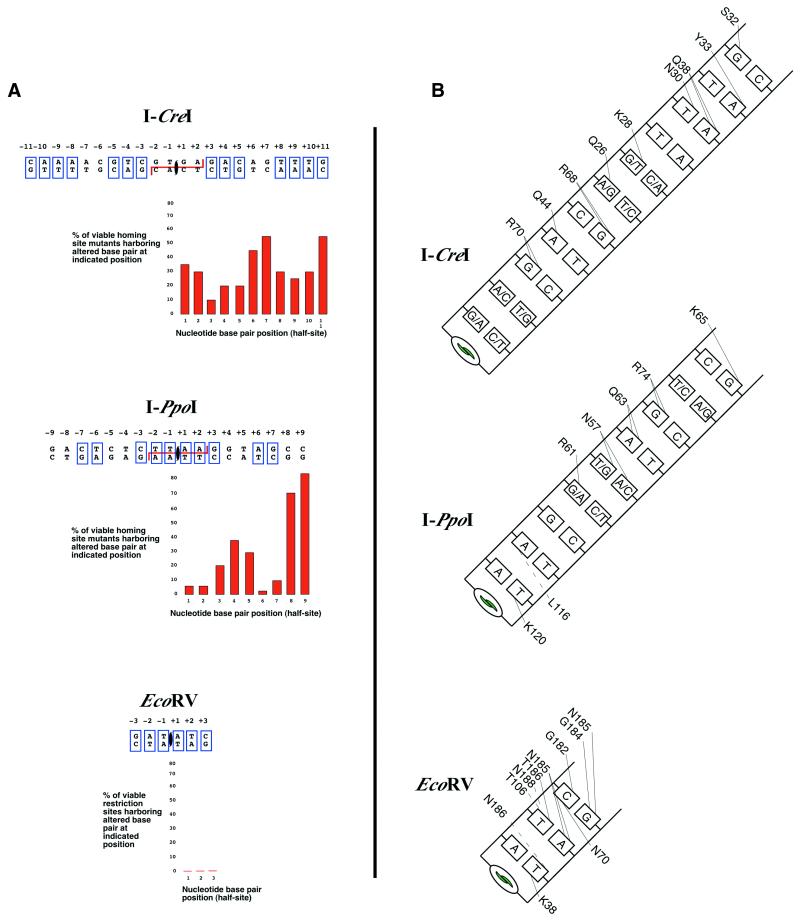
DNA-binding by homing and
restriction endonucleases. (A) Summary of actively
cleaved target sites containing single base changes. I-CreI
and I-PpoI recognize degenerate palindromes; EcoRV recognizes a strict palindrome. Palindromic
bases are boxed. The homing endonucleases I-CreI
and I-PpoI are tolerant of minor changes in their
homing site as shown. The restriction endonuclease EcoRV
is intolerant of any changes to its restriction site. Generally,
the level of tolerance correlates inversely to number of specific
protein–DNA contacts. Mutant homing site data taken from
Argast et al. (114)
and restriction site data from Pingoud and Jeltsch (112).
(B) Base-specific contacts made by each endonuclease.
Major groove contacts are drawn to the top of the base; minor groove
contacts are drawn to the bottom of the base. Hydrogen bonds are
represented as solid lines; other interactions are dashed lines.
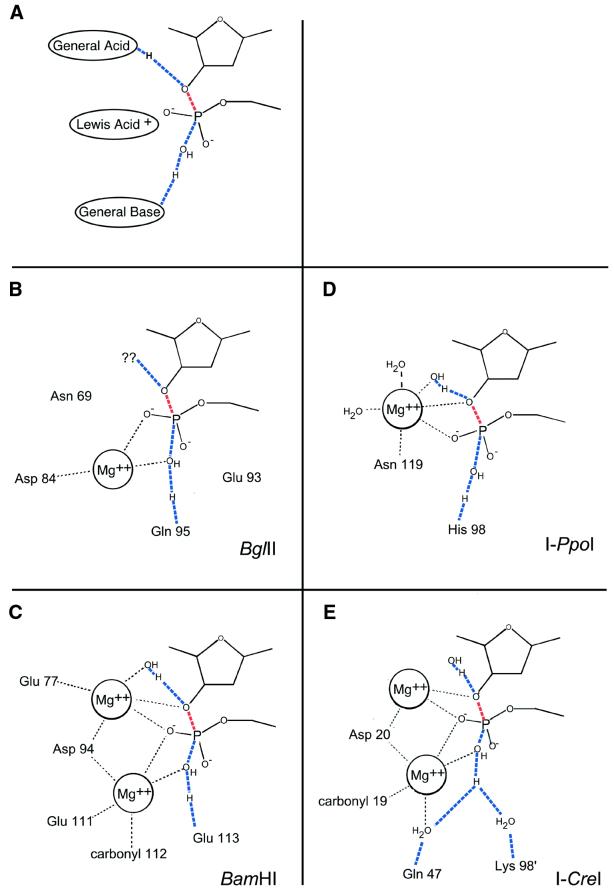
Endonuclease cleavage transition
states. In all figures, the blue lines represent bonds in transition.
Red lines represent the breaking bond of the DNA backbone. (A)
Phosphodiester bond cleavage requires three chemical entities: a
general base to activate the nucleophile, a Lewis acid to stabilize
the pentacoordinate phosphoanion transition state and a general
acid to protonate the 3′ leaving group.
Water molecules usually function as nucleophiles and often as general
acids. Divalent metal cations function as Lewis acids, can help
lower the pKa of associated waters and
position the nucleophile. Other residues act as general bases and/or to position
waters and metal ions. (B) BglII
uses a single metal in its mechanism. The magnesium functions to
lower pKa of nucleophile and stabilize
the transition state. The general base appears to be Gln95; the
general acid has not been conclusively identified. (C)
In BamHI, two magnesium ions aid in catalysis by
positioning and lowering the pKa of
the nucleophile and proton donor and stabilizing the phosphoanion
transition state. Glu113 is the general base and water is the general
acid. (D) I-PpoI, like BglII,
uses a single-metal mechanism. In I-PpoI, the metal
stabilizes the transition state and activates the general acid,
an associated water molecule. It does not appear to participate
in positioning and/or activating the nucleophile. The general
base is His98. (E) I-CreI uses
a two-metal mechanism, the central metal being shared by two separate
active sites. The metals position and lower the pKa of
the nucleophile, stabilize the transition state and stabilize the 3′ oxygen leaving group. The
general base and acid appear to be within a well-ordered solvent
network (positioned by peripheral residues and the first metal ion); for
clarity, only three of these water molecules—other than
the nucleophile—are drawn.

Structural alignment of LAGLIDADG
active sites. View is top-down through protein body to active site
residues and DNA below. In all three diagrams, subunits of the homodimer
I-CreI are blue and purple and are labeled as I-CreI and I-CreI′.
The DNA backbone (gray) and the three divalent ions (black) from
I-CreI–DNA co-crystal structures are shown
for reference. Note the enormous divergence in both amino acid identity
and position at the periphery of the active sites. (A)
I-DmoI over I-CreI. (B)
PI-PfuI over I-CreI. (C)
PI-SceI over I-CreI. Alignments
generated by superposition of LAGLIDADG α-helices.

Diagram of the I-CreI
active site (A) in the presence of calcium (non-cleaved
DNA substrate) and (B) in the presence of magnesium
(cleaved DNA substrate). DNA is red, metal ions are green, protein
side chains are black. Waters are light blue; distances between
waters are shown in Å by the associated blue lines representing
the hydrogen-bonding network. The nucleophilic water is purple.
Note that the protein makes no direct contact to the nucleophile,
the scissile phosphate (red outlined in black), or the 3′ leaving
group and that the solvent network extends from the nucleophile
to the leaving group.
References
-
- Dujon B., Belfort,M., Butow,R.A., Jacq,C., Lemieux,C., Perlman,P.S. and Vogt,V.M. (1989) Mobile introns: definition of terms and recommended nomenclature. Gene, 82, 115–118. - PubMed
-
- Coen D., Deutsch,J., Netter,P., Petrochilo,E. and Slonimski,P.P. (1970) Mitochondrial genetics. I. Methodology and phenomenology. Symp. Soc. Exp. Biol., 23, 449–496. - PubMed
-
- Dujon B. (1980) Sequence of the intron and flanking exons of the mitochondrial 21S rRNA gene of yeast strains having different alleles at the omega and rib-1 loci. Cell, 20, 185–197. - PubMed
-
- Bos J.L., Heyting,C., Borst,P., Arnberg,A.C. and Van Bruggen,E.F. (1978) An insert in the single gene for the large ribosomal RNA in yeast mitochondrial DNA. Nature, 275, 336–338. - PubMed
-
- Colleaux L., d’Auriol,L., Betermier,M., Cottarel,G., Jacquier,A., Galibert,F. and Dujon,B. (1986) Universal code equivalent of a yeast mitochondrial intron reading frame is expressed into E.coli as a specific double strand endonuclease. Cell, 44, 521–533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
