

Novel critical role of a human Mediator complex for basal RNA polymerase II transcription
- PMID: 11559591
- PMCID: PMC1084041
- DOI: 10.1093/embo-reports/kve186
Novel critical role of a human Mediator complex for basal RNA polymerase II transcription
Abstract
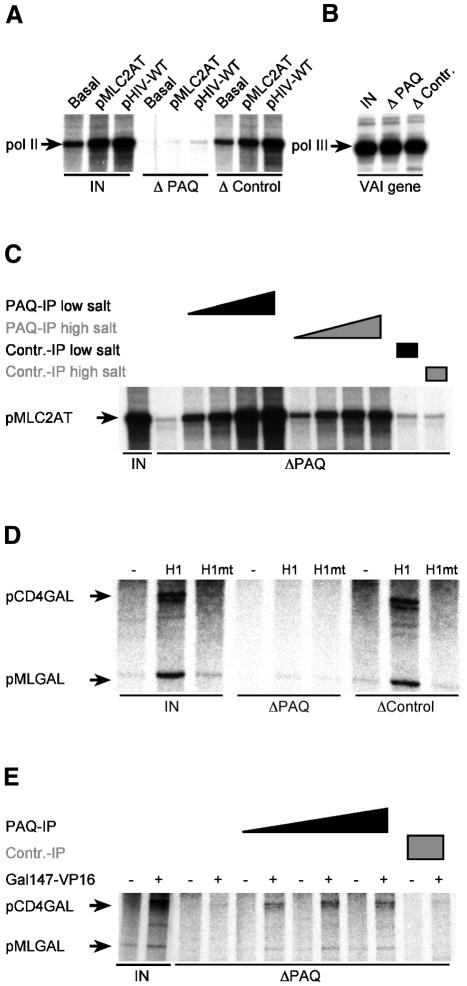
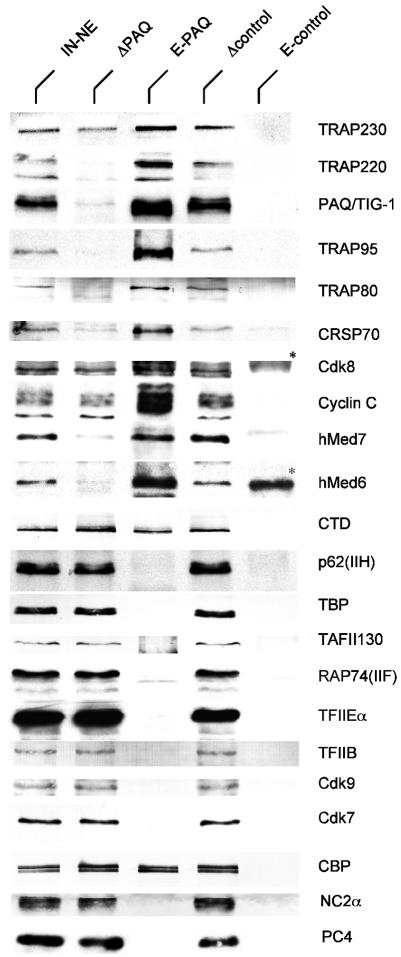
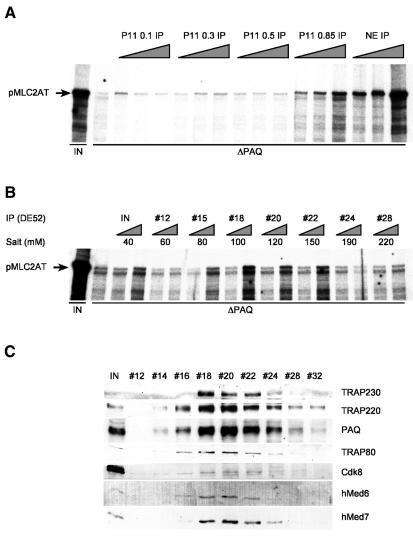
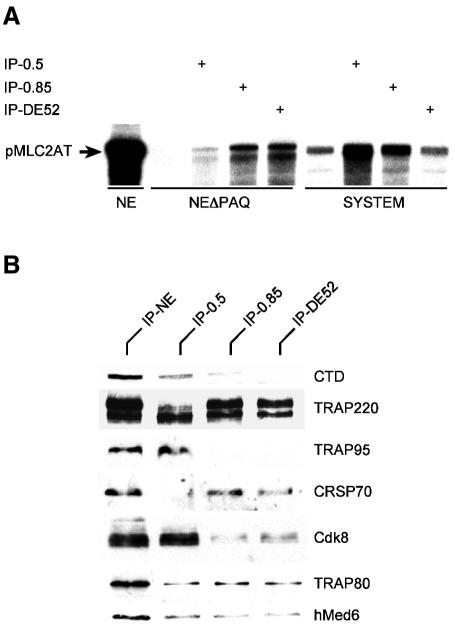
Human Mediator complexes have been described as important bridging factors that enhance the effect of activators in purified systems and in chromatin. Here we report a novel basal function of a human Mediator complex. A monoclonal antibody was generated that depleted the majority of Mediator components from crude cell extracts. The removal of human Mediator abolished transcription by RNA polymerase II. This was observed on all genes tested, on TATA-containing and TATA-less promoters, both in the presence and absence of activators. To identify the relevant complex a combined biochemical and immunopurification protocol was applied. Two variants termed Mediator and basal Mediator were functionally and structurally distinguished. Basal Mediator function relies on additional constraints, which is reflected in the observation that it is essential in crude but not in purified systems. We conclude that basal Mediator is a novel general transcription factor of RNA polymerase II.
Figures
References
-
- Abraham S. and Solomon, W.B. (2000) A novel glutamine-rich putative transcriptional adaptor protein (TIG-1), preferentially expressed in placental and bone-marrow tissues. Gene, 255, 389–400. - PubMed
-
- Akoulitchev S., Chuikov, S. and Reinberg, D. (2000) TFIIH is negatively regulated by cdk8-containing mediator complexes. Nature, 407, 102–106. - PubMed
-
- Berti L. et al. (2001) Isolation and characterization of a novel gene from the DiGeorge chromosomal region that encodes for a Mediator subunit. Genomics, 74, 320–332. - PubMed
-
- Boyer T.G., Martin, M.E., Lees, E., Ricciardi, R.P. and Berk, A.J. (1999) Mammalian Srb/Mediator complex is targeted by adenovirus E1A protein. Nature, 399, 276–279. - PubMed
-
- Flanagan P.M., Kelleher, R.J., Sayre, M.H., Tschochner, H. and Kornberg, R.D. (1991) A mediator required for activation of RNA polymerase II transcription in vitro. Nature, 350, 436–438. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
