

Actin pedestal formation by enteropathogenic Escherichia coli and intracellular motility of Shigella flexneri are abolished in N-WASP-defective cells
- PMID: 11559594
- PMCID: PMC1084051
- DOI: 10.1093/embo-reports/kve197
Actin pedestal formation by enteropathogenic Escherichia coli and intracellular motility of Shigella flexneri are abolished in N-WASP-defective cells
Abstract
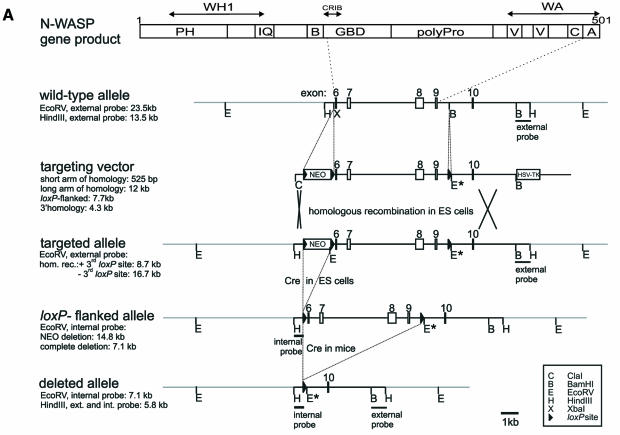
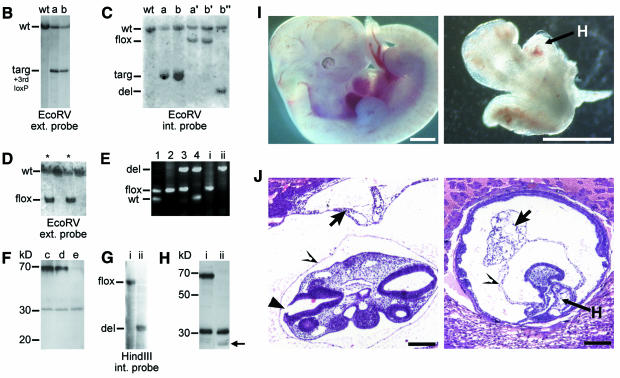
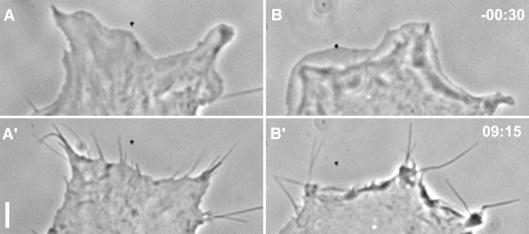
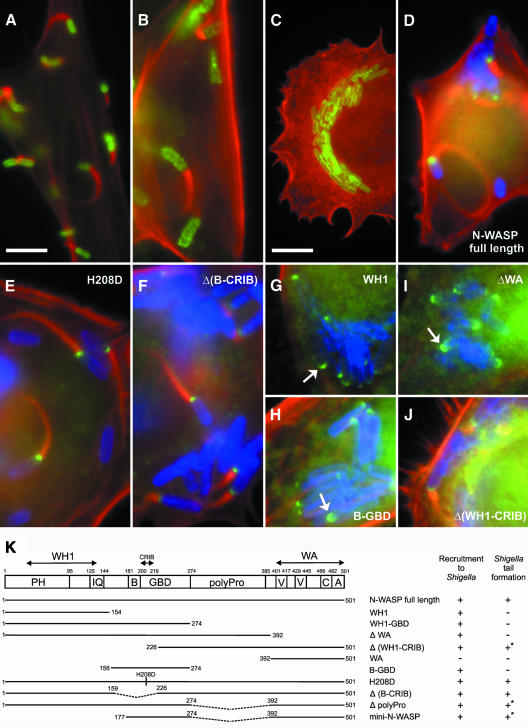
In mammalian cells, actin dynamics is tightly controlled through small GTPases of the Rho family, WASP/Scar proteins and the Arp2/3 complex. We employed Cre/loxP-mediated gene targeting to disrupt the ubiquitously expressed N-WASP in the mouse germline, which led to embryonic lethality. To elucidate the role of N-WASP at the cellular level, we immortalized embryonic fibroblasts and selected various N-WASP-defective cell lines. These fibroblasts showed no apparent morphological alterations and were highly responsive to the induction of filopodia, but failed to support the motility of Shigella flexneri. In addition, enteropathogenic Escherichia coli were incapable of inducing the formation of actin pedestals in N-WASP-defective cells. Our results prove the essential role of this protein for actin cytoskeletal changes induced by these bacterial pathogens in vivo and in addition show for the first time that N-WASP is dispensable for filopodia formation.
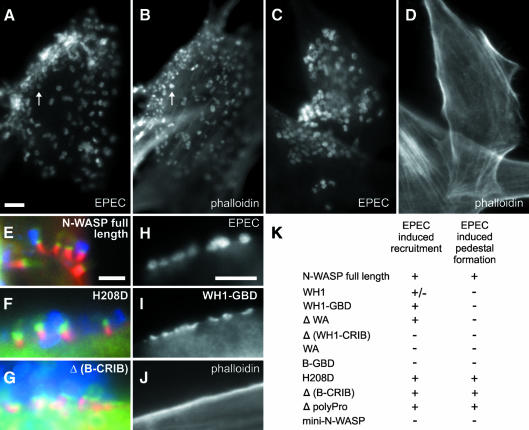
Figures
Similar articles
-
Activation of the CDC42 effector N-WASP by the Shigella flexneri IcsA protein promotes actin nucleation by Arp2/3 complex and bacterial actin-based motility.J Cell Biol. 1999 Sep 20;146(6):1319-32. doi: 10.1083/jcb.146.6.1319. J Cell Biol. 1999. PMID: 10491394 Free PMC article.
-
Rho family GTPase Cdc42 is essential for the actin-based motility of Shigella in mammalian cells.J Exp Med. 2000 Jun 5;191(11):1905-20. doi: 10.1084/jem.191.11.1905. J Exp Med. 2000. PMID: 10839806 Free PMC article.
-
WIP regulates N-WASP-mediated actin polymerization and filopodium formation.Nat Cell Biol. 2001 May;3(5):484-91. doi: 10.1038/35074551. Nat Cell Biol. 2001. PMID: 11331876
-
Signalling to actin: the Cdc42-N-WASP-Arp2/3 connection.Chem Biol. 1999 Sep;6(9):R235-40. doi: 10.1016/s1074-5521(99)80107-0. Chem Biol. 1999. PMID: 10467124 Review.
-
Wiskott-Aldrich syndrome protein, WASP.Int J Biochem Cell Biol. 1999 Mar-Apr;31(3-4):383-7. doi: 10.1016/s1357-2725(98)00118-6. Int J Biochem Cell Biol. 1999. PMID: 10224664 Review.
Cited by
-
Enteropathogenic Escherichia coli and vaccinia virus do not require the family of WASP-interacting proteins for pathogen-induced actin assembly.Infect Immun. 2012 Dec;80(12):4071-7. doi: 10.1128/IAI.06148-11. Epub 2012 Sep 10. Infect Immun. 2012. PMID: 22966049 Free PMC article.
-
Fusion-Independent Satellite Cell Communication to Muscle Fibers During Load-Induced Hypertrophy.Function (Oxf). 2020;1(1):zqaa009. doi: 10.1093/function/zqaa009. Epub 2020 Jul 6. Function (Oxf). 2020. PMID: 32864621 Free PMC article.
-
Effects of ectopically expressed neuronal Wiskott-Aldrich syndrome protein domains on Rickettsia rickettsii actin-based motility.Infect Immun. 2003 Mar;71(3):1551-6. doi: 10.1128/IAI.71.3.1551-1556.2003. Infect Immun. 2003. PMID: 12595475 Free PMC article.
-
The ability of an attaching and effacing pathogen to trigger localized actin assembly contributes to virulence by promoting mucosal attachment.Cell Microbiol. 2014 Sep;16(9):1405-24. doi: 10.1111/cmi.12302. Epub 2014 Jun 2. Cell Microbiol. 2014. PMID: 24780054 Free PMC article.
-
Conditional knock out of N-WASP in keratinocytes causes skin barrier defects and atopic dermatitis-like inflammation.Sci Rep. 2017 Aug 4;7(1):7311. doi: 10.1038/s41598-017-07125-8. Sci Rep. 2017. PMID: 28779153 Free PMC article.
References
-
- Aronheim A., Broder, Y.C., Cohen, A., Fritsch, A., Belisle, B. and Abo, A. (1998) Chp, a homologue of the GTPase Cdc42Hs, activates the JNK pathway and is implicated in reorganizing the actin cytoskeleton. Curr. Biol., 8, 1125–1128. - PubMed
-
- Aspenstrom P., Lindberg, U. and Hall, A. (1996) Two GTPases, Cdc42 and Rac, bind directly to a protein implicated in the immunodeficiency disorder Wiskott–Aldrich syndrome. Curr. Biol., 6, 70–75. - PubMed
-
- Carl U.D., Pollmann, M., Orr, E., Gertler, F.B., Chakraborty, T. and Wehland, J. (1999) Aromatic and basic residues within the EVH1 domain of VASP specify its interaction with proline-rich ligands. Curr. Biol., 9, 715–718. - PubMed
-
- Carlier M.F., Ducruix, A. and Pantaloni, D. (1999) Signalling to actin: the Cdc42-N-WASP-Arp2/3 connection. Chem. Biol., 6, R235–R240. - PubMed
-
- Carlier M.F. et al. (2000) GRB2 links signaling to actin assembly by enhancing interaction of neural Wiskott–Aldrich syndrome protein (N-WASp) with actin-related protein (ARP2/3) complex. J. Biol. Chem., 275, 21946–21952. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
