

A post-docking role for active zone protein Rim
- PMID: 11559854
- PMCID: PMC2585766
- DOI: 10.1038/nn732
A post-docking role for active zone protein Rim
Abstract
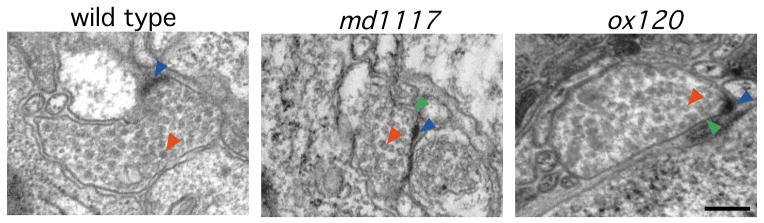
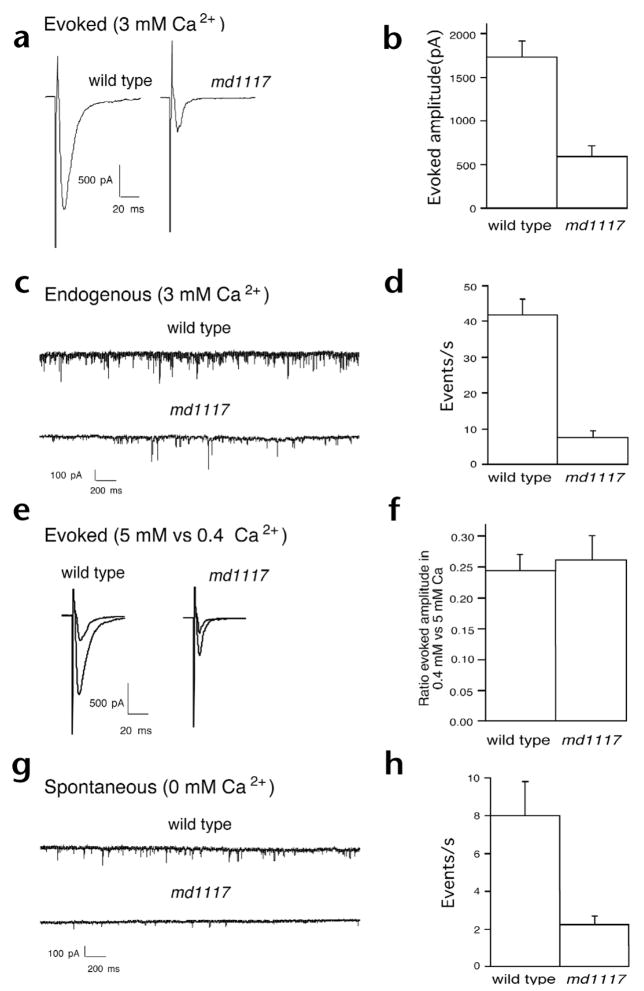
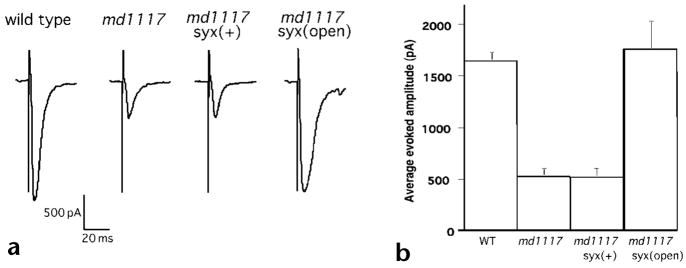
Rim1 was previously identified as a Rab3 effector localized to the presynaptic active zone in vertebrates. Here we demonstrate that C. elegans unc-10 mutants lacking Rim are viable, but exhibit behavioral and physiological defects that are more severe than those of Rab3 mutants. Rim is localized to synaptic sites in C. elegans, but the ultrastructure of the presynaptic densities is normal in Rim mutants. Moreover, normal levels of docked synaptic vesicles were observed in mutants, suggesting that Rim is not involved in the docking process. The level of fusion competent vesicles at release sites was reduced fivefold in Rim mutants, but calcium sensitivity of release events was unchanged. Furthermore, expression of a constitutively open form of syntaxin suppressed the physiological defects of Rim mutants, suggesting Rim normally acts to regulate conformational changes in syntaxin. These data suggest Rim acts after vesicle docking likely via regulating priming.
Figures
Comment in
-
pRIMing synaptic vesicles for fusion.Nat Neurosci. 2001 Oct;4(10):965-6. doi: 10.1038/nn1001-965. Nat Neurosci. 2001. PMID: 11574826 No abstract available.
Similar articles
-
Rabphilin potentiates soluble N-ethylmaleimide sensitive factor attachment protein receptor function independently of rab3.J Neurosci. 2001 Dec 1;21(23):9255-64. doi: 10.1523/JNEUROSCI.21-23-09255.2001. J Neurosci. 2001. PMID: 11717359 Free PMC article.
-
Direct interactions between C. elegans RAB-3 and Rim provide a mechanism to target vesicles to the presynaptic density.Neurosci Lett. 2008 Oct 24;444(2):137-42. doi: 10.1016/j.neulet.2008.08.026. Epub 2008 Aug 14. Neurosci Lett. 2008. PMID: 18721860 Free PMC article.
-
Rim1 and rabphilin-3 bind Rab3-GTP by composite determinants partially related through N-terminal alpha -helix motifs.J Biol Chem. 2001 Aug 31;276(35):32480-8. doi: 10.1074/jbc.M103337200. Epub 2001 Jun 28. J Biol Chem. 2001. PMID: 11431472
-
The synaptic vesicle cycle: exocytosis and endocytosis in Drosophila and C. elegans.Curr Opin Neurobiol. 2002 Oct;12(5):499-507. doi: 10.1016/s0959-4388(02)00360-4. Curr Opin Neurobiol. 2002. PMID: 12367628 Review.
-
Synaptic vesicle docking: a putative role for the Munc18/Sec1 protein family.Curr Top Dev Biol. 2005;65:83-113. doi: 10.1016/S0070-2153(04)65003-4. Curr Top Dev Biol. 2005. PMID: 15642380 Review. No abstract available.
Cited by
-
Munc18-1 binds directly to the neuronal SNARE complex.Proc Natl Acad Sci U S A. 2007 Feb 20;104(8):2697-702. doi: 10.1073/pnas.0611318104. Epub 2007 Feb 14. Proc Natl Acad Sci U S A. 2007. PMID: 17301226 Free PMC article.
-
RIM is essential for stimulated but not spontaneous somatodendritic dopamine release in the midbrain.Elife. 2019 Sep 5;8:e47972. doi: 10.7554/eLife.47972. Elife. 2019. PMID: 31486769 Free PMC article.
-
Redundant localization mechanisms of RIM and ELKS in Caenorhabditis elegans.J Neurosci. 2005 Jun 22;25(25):5975-83. doi: 10.1523/JNEUROSCI.0804-05.2005. J Neurosci. 2005. PMID: 15976086 Free PMC article.
-
SNARE assembly enlightened by cryo-EM structures of a synaptobrevin-Munc18-1-syntaxin-1 complex.Sci Adv. 2022 Jun 24;8(25):eabo5272. doi: 10.1126/sciadv.abo5272. Epub 2022 Jun 22. Sci Adv. 2022. PMID: 35731863 Free PMC article.
-
Presynaptic Calcium Channels.Int J Mol Sci. 2019 May 6;20(9):2217. doi: 10.3390/ijms20092217. Int J Mol Sci. 2019. PMID: 31064106 Free PMC article. Review.
References
-
- Burns M, Augustine G. Synaptic structure and function: dynamic organization yields architectural precision. Cell. 1995;83:187–194. - PubMed
-
- Heuser JE, Reese TS. In: Handbook of Physiology I: The Nervous System. Kandel ER, editor. American Physiological Society: Baltimore; 1973. pp. 261–294.
-
- Landis DM, Hall AK, Weinstein LA, Reese TS. The organization of cytoplasm at the presynaptic active zone of a central nervous system synapse. Neuron. 1988;1:201–209. - PubMed
-
- Garner CC, Kindler S, Gundelfinger ED. Molecular determinants of presynaptic active zones. Curr Opin Neurobiol. 2000;10:321–327. - PubMed
-
- Wang Y, Okamoto M, Schmitz F, Hofmann K, Südhof TC. Rim is a putative Rab3 effector in regulating synaptic-vesicle fusion. Nature. 1997;388:593–598. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
