

Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo
- PMID: 11560993
- PMCID: PMC2195961
- DOI: 10.1084/jem.194.6.769
Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo
Abstract
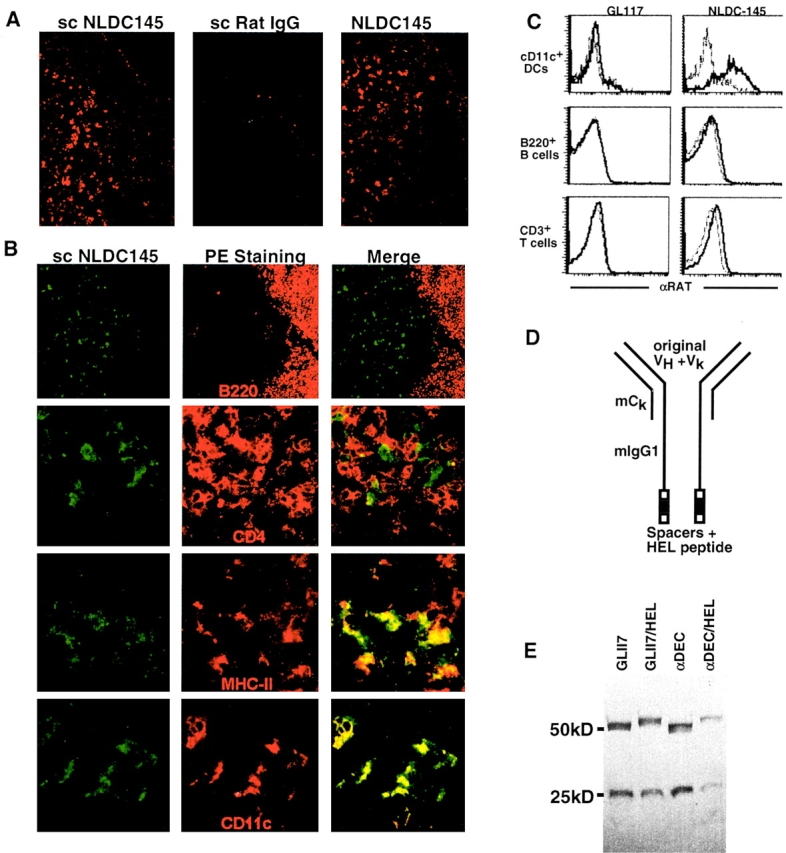
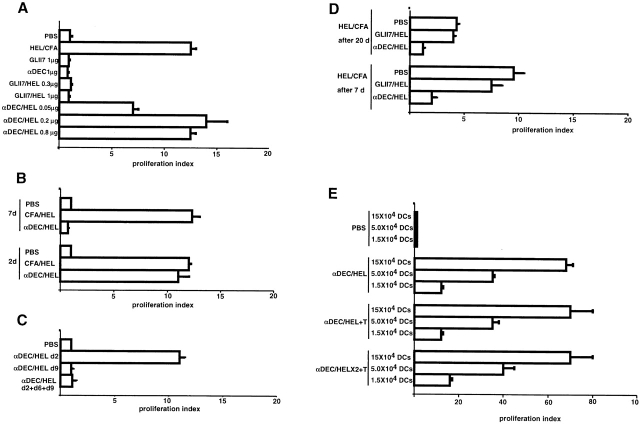
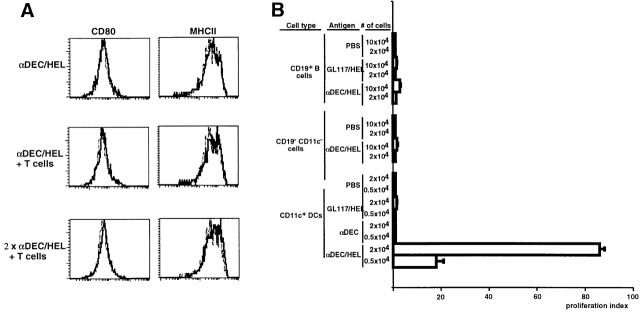
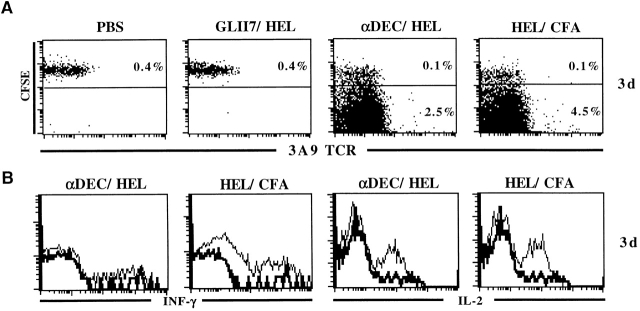
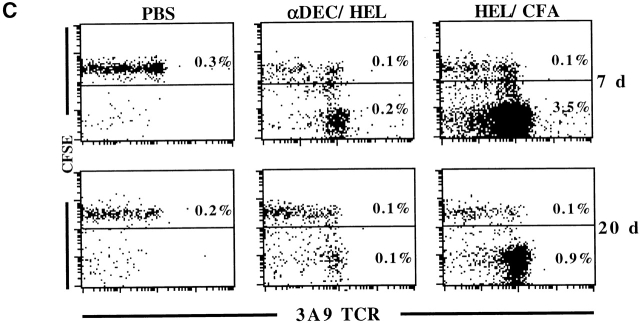
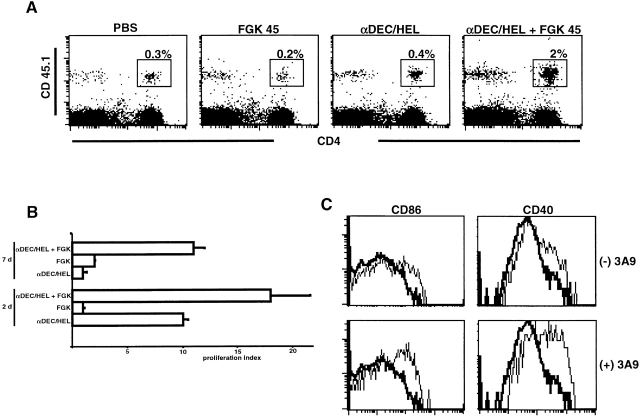
Dendritic cells (DCs) have the capacity to initiate immune responses, but it has been postulated that they may also be involved in inducing peripheral tolerance. To examine the function of DCs in the steady state we devised an antigen delivery system targeting these specialized antigen presenting cells in vivo using a monoclonal antibody to a DC-restricted endocytic receptor, DEC-205. Our experiments show that this route of antigen delivery to DCs is several orders of magnitude more efficient than free peptide in complete Freund's adjuvant (CFA) in inducing T cell activation and cell division. However, T cells activated by antigen delivered to DCs are not polarized to produce T helper type 1 cytokine interferon gamma and the activation response is not sustained. Within 7 d the number of antigen-specific T cells is severely reduced, and the residual T cells become unresponsive to systemic challenge with antigen in CFA. Coinjection of the DC-targeted antigen and anti-CD40 agonistic antibody changes the outcome from tolerance to prolonged T cell activation and immunity. We conclude that in the absence of additional stimuli DCs induce transient antigen-specific T cell activation followed by T cell deletion and unresponsiveness.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
