

Membrane raft-dependent regulation of phospholipase Cgamma-1 activation in T lymphocytes
- PMID: 11564877
- PMCID: PMC99870
- DOI: 10.1128/MCB.21.20.6939-6950.2001
Membrane raft-dependent regulation of phospholipase Cgamma-1 activation in T lymphocytes
Abstract
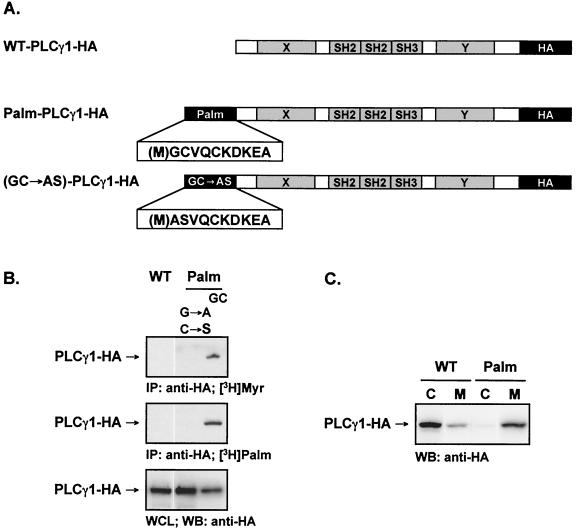
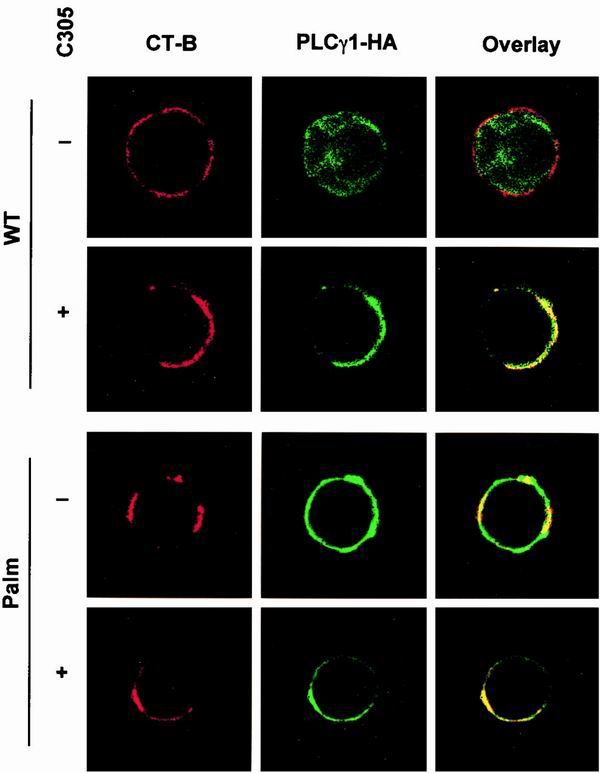
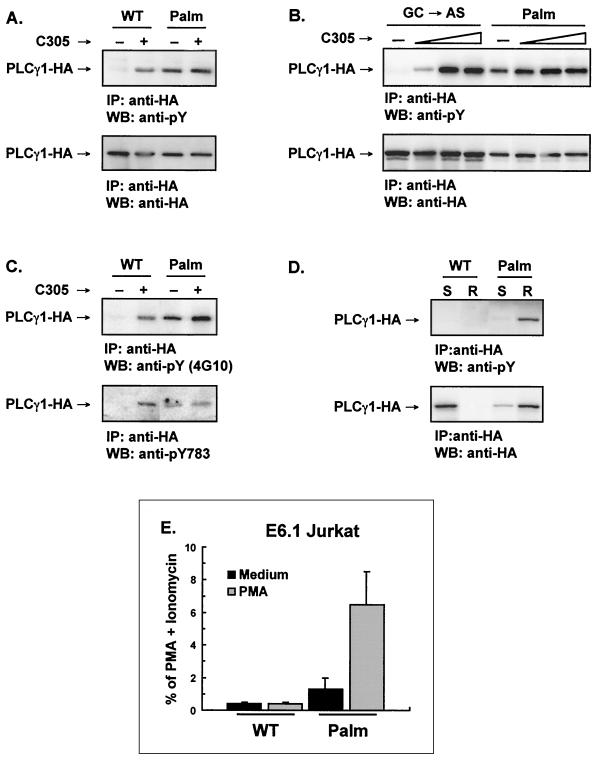
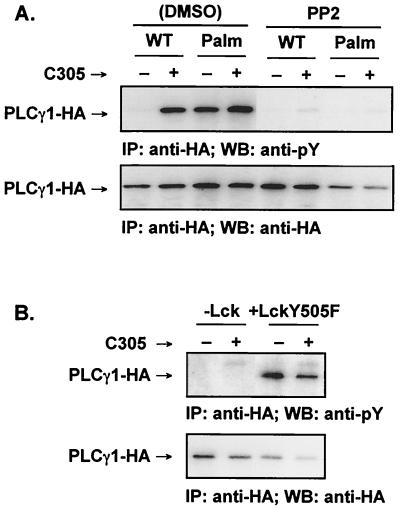
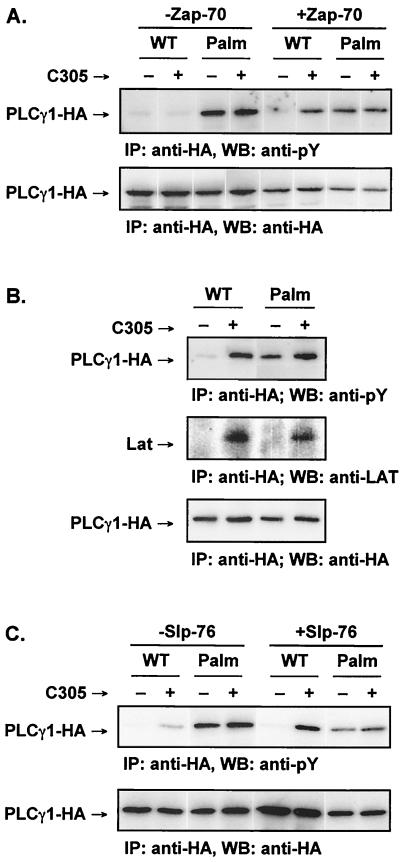
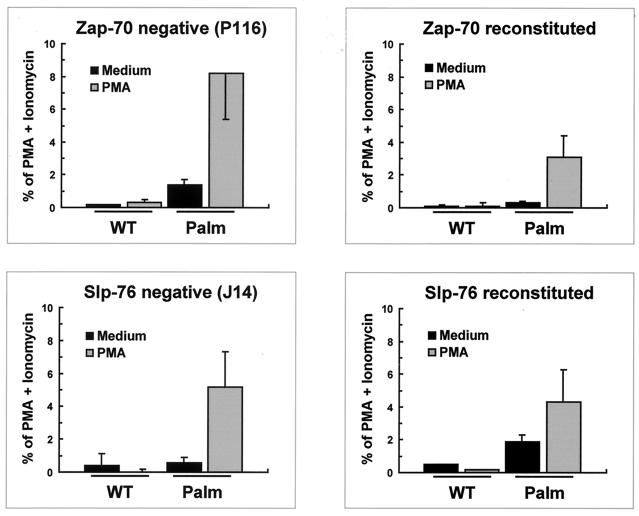
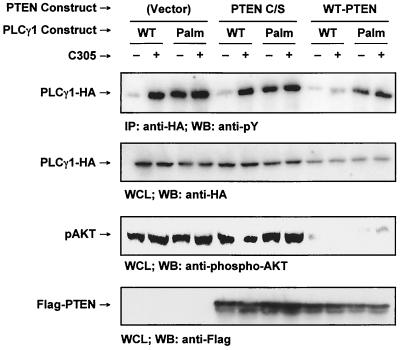
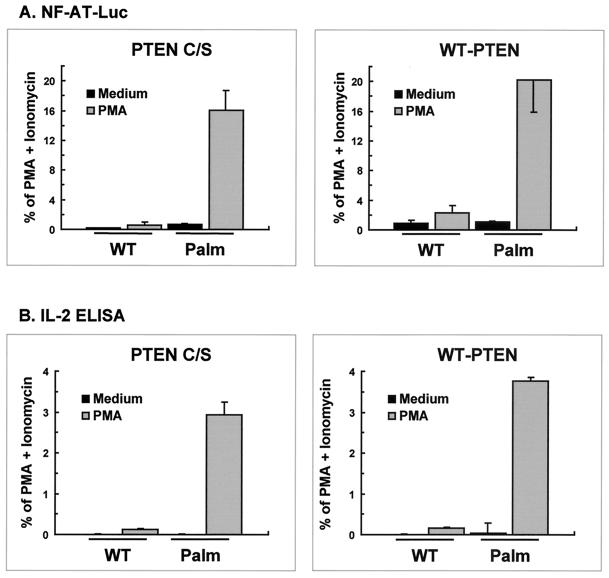
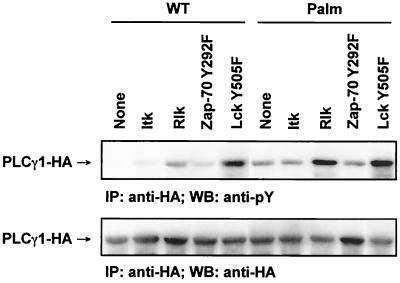
Numerous signaling molecules associate with lipid rafts, either constitutively or after engagement of surface receptors. One such molecule, phospholipase Cgamma-1 (PLCgamma1), translocates from the cytosol to lipid rafts during T-cell receptor (TCR) signaling. To investigate the role played by lipid rafts in the activation of this molecule in T cells, an influenza virus hemagglutinin A (HA)-tagged PLCgamma1 was ectopically expressed in Jurkat T cells and targeted to these microdomains by the addition of a dual-acylation signal. Raft-targeted PLCgamma1 was constitutively tyrosine phosphorylated and induced constitutive NF-AT-dependent transcription and interleukin-2 secretion in Jurkat cells. Tyrosine phosphorylation of raft-targeted PLCgamma1 did not require Zap-70 or the interaction with the adapters Lat and Slp-76, molecules that are necessary for TCR signaling. In contrast, the Src family kinase Lck was required. Coexpression in HEK 293T cells of PLCgamma1-HA with Lck or the Tec family kinase Rlk resulted in preferential phosphorylation of raft-targeted PLCgamma1 over wild-type PLCgamma1. These data show that localization of PLCgamma1 in lipid rafts is sufficient for its activation and demonstrate a role for lipid rafts as microdomains that dynamically segregate and integrate PLCgamma1 with other signaling components.
Figures

References
-
- Brown D A, London E. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J Biol Chem. 2000;275:17221–17224. - PubMed
-
- Crabtree G R, Clipstone N A. Signal transmission between the plasma membrane and nucleus of T lymphocytes. Annu Rev Biochem. 1994;63:1045–1083. - PubMed
-
- DeBell K E, Stoica B A, Veri M C, Di Baldassarre A, Miscia S, Graham L J, Rellahan B L, Ishiai M, Kurosaki T, Bonvini E. Functional independence and interdependence of the src homology domains of phospholipase C-gamma1 in B-cell receptor signal transduction. Mol Cell Biol. 1999;19:7388–7398. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous