

Mitochondrial import driving forces: enhanced trapping by matrix Hsp70 stimulates translocation and reduces the membrane potential dependence of loosely folded preproteins
- PMID: 11564892
- PMCID: PMC99885
- DOI: 10.1128/MCB.21.20.7097-7104.2001
Mitochondrial import driving forces: enhanced trapping by matrix Hsp70 stimulates translocation and reduces the membrane potential dependence of loosely folded preproteins
Abstract
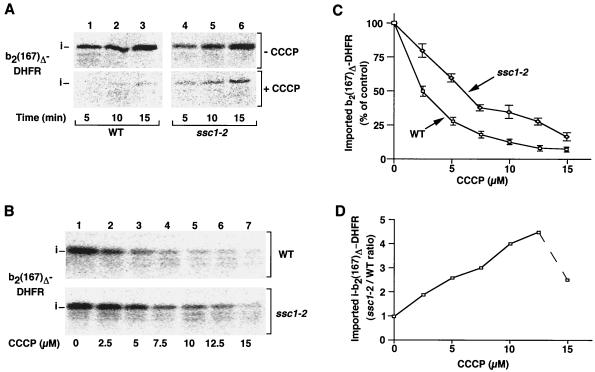
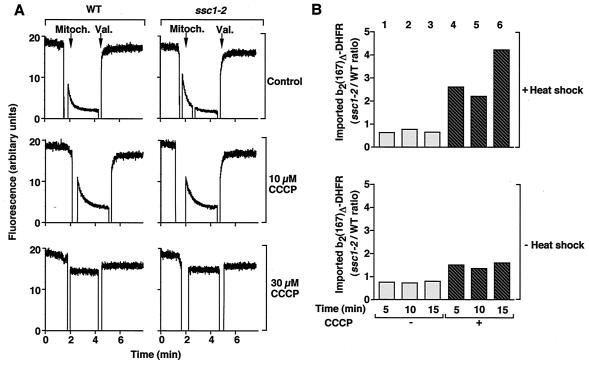
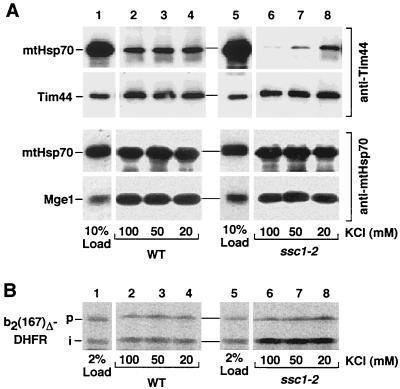
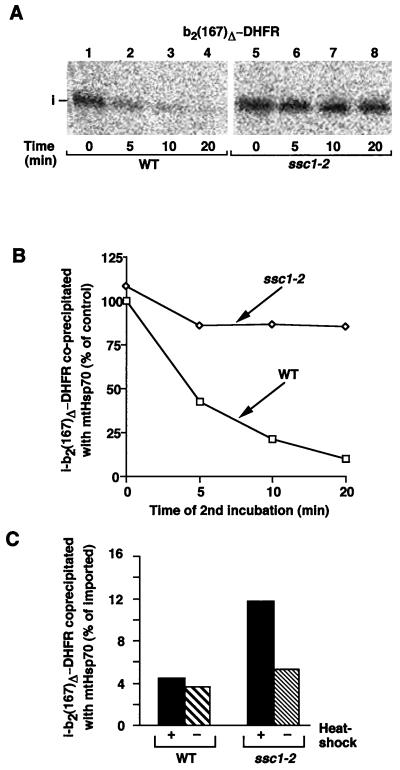
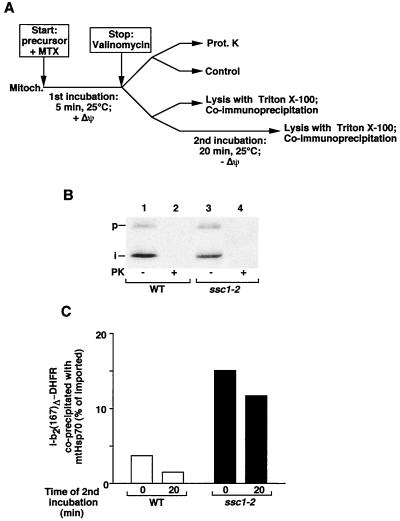
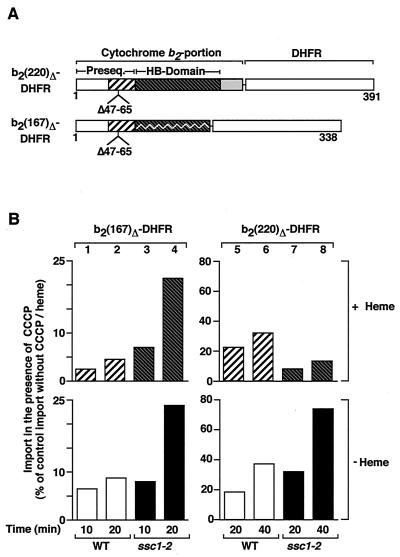
The mitochondrial heat shock protein Hsp70 (mtHsp70) is essential for driving translocation of preproteins into the matrix. Two models, trapping and pulling by mtHsp70, are discussed, but positive evidence for either model has not been found so far. We have analyzed a mutant mtHsp70, Ssc1-2, that shows a reduced interaction with the membrane anchor Tim44, but an enhanced trapping of preproteins. Unexpectedly, at a low inner membrane potential, ssc1-2 mitochondria imported loosely folded preproteins more efficiently than wild-type mitochondria. The import of a tightly folded preprotein, however, was not increased in ssc1-2 mitochondria. Thus, enhanced trapping by mtHsp70 stimulates the import of loosely folded preproteins and reduces the dependence on the import-driving activity of the membrane potential, directly demonstrating that trapping is one of the molecular mechanisms of mtHsp70 action.
Figures
References
-
- Alconada A, Gärtner F, Hönlinger A, Kübrich M, Pfanner N. Mitochondrial receptor complex from Neurospora crassa and Saccharomyces cerevisiae. Methods Enzymol. 1995;260:263–286. - PubMed
-
- Bauer M F, Hofmann S, Neupert W, Brunner M. Protein translocation into mitochondria: the role of TIM complexes. Trends Cell Biol. 2000;10:25–31. - PubMed
-
- Bauer M F, Sirrenberg C, Neupert W, Brunner M. Role of Tim23 as voltage sensor and presequence receptor in protein import into mitochondria. Cell. 1996;87:33–41. - PubMed
-
- Bömer U, Maarse A, Martin F, Geissler A, Merlin A, Schonfisch B, Meijer M, Pfanner N, Rassow J. Separation of structural and dynamic functions of the mitochondrial translocase: Tim44 is crucial for the inner membrane import sites in translocation of tightly folded domains, but not of loosely folded preproteins. EMBO J. 1998;17:4226–4237. - PMC - PubMed
-
- Bukau B, Horwich A. The Hsp70 and Hsp60 chaperone machines. Cell. 1998;92:351–366. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases