

Citrin and aralar1 are Ca(2+)-stimulated aspartate/glutamate transporters in mitochondria
- PMID: 11566871
- PMCID: PMC125626
- DOI: 10.1093/emboj/20.18.5060
Citrin and aralar1 are Ca(2+)-stimulated aspartate/glutamate transporters in mitochondria
Abstract
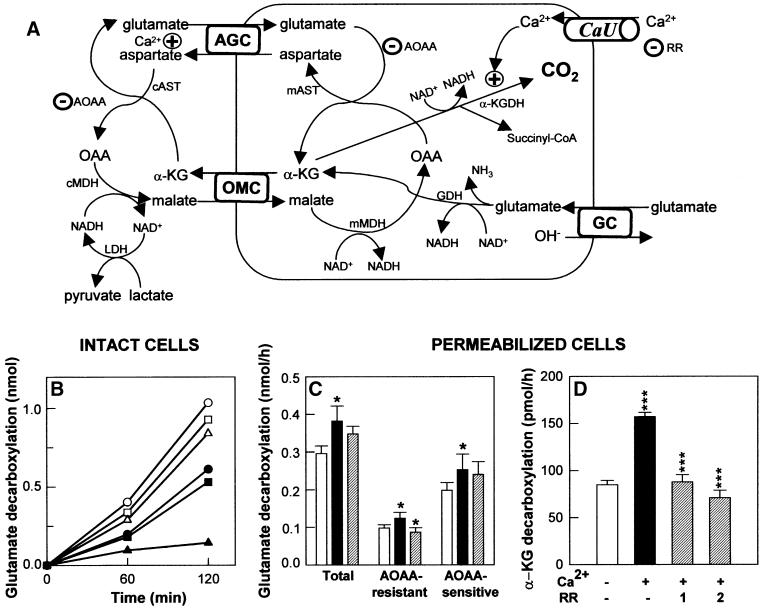
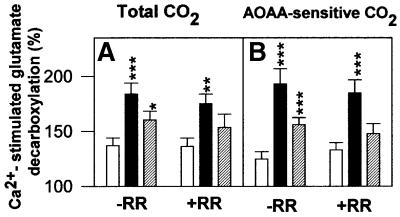
The mitochondrial aspartate/glutamate carrier catalyzes an important step in both the urea cycle and the aspartate/malate NADH shuttle. Citrin and aralar1 are homologous proteins belonging to the mitochondrial carrier family with EF-hand Ca(2+)-binding motifs in their N-terminal domains. Both proteins and their C-terminal domains were overexpressed in Escherichia coli, reconstituted into liposomes and shown to catalyze the electrogenic exchange of aspartate for glutamate and a H(+). Overexpression of the carriers in transfected human cells increased the activity of the malate/aspartate NADH shuttle. These results demonstrate that citrin and aralar1 are isoforms of the hitherto unidentified aspartate/glutamate carrier and explain why mutations in citrin cause type II citrullinemia in humans. The activity of citrin and aralar1 as aspartate/glutamate exchangers was stimulated by Ca(2+) on the external side of the inner mitochondrial membrane, where the Ca(2+)-binding domains of these proteins are localized. These results show that the aspartate/glutamate carrier is regulated by Ca(2+) through a mechanism independent of Ca(2+) entry into mitochondria, and suggest a novel mechanism of Ca(2+) regulation of the aspartate/malate shuttle.
Figures
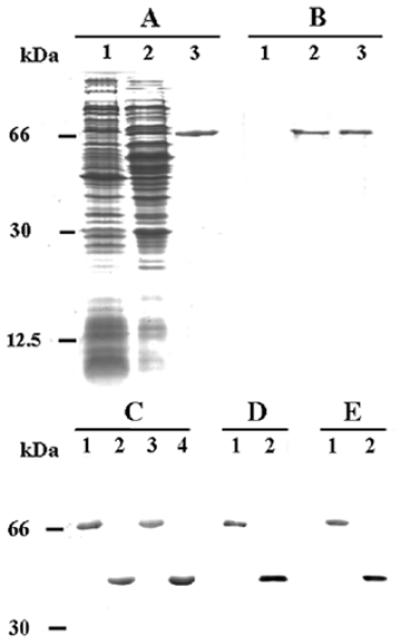
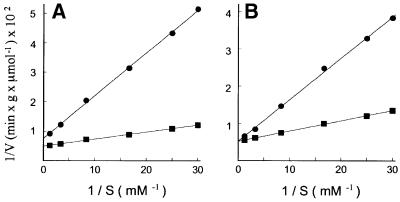
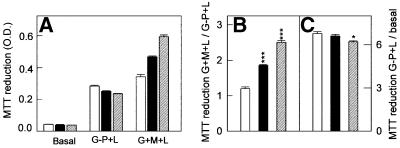
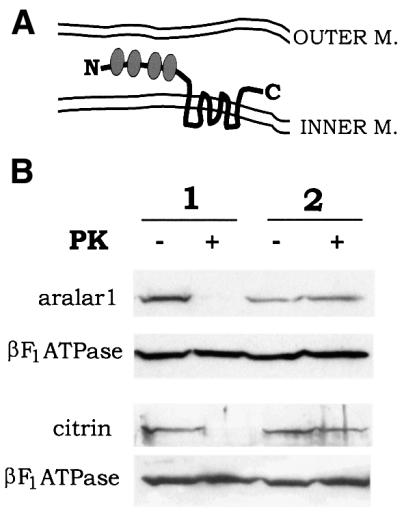
References
-
- Berridge M.V. and Tan,A.S. (1993) Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys., 303, 474–482. - PubMed
-
- Bisaccia F., De Palma,A. and Palmieri,F. (1992) Identification and purification of the aspartate/glutamate carrier from bovine heart mitochondria. Biochim. Biophys. Acta, 1106, 291–296. - PubMed
-
- Brown L.J., MacDonald,M.J., Lehn,D.A. and Moran,S.M. (1994) Sequence of rat mitochondrial glycerol-3-phosphate dehydrogenase cDNA. J. Biol. Chem., 269, 14363–14366. - PubMed
-
- Bunger R. and Mallet,R.T. (1993) Mitochondrial pyruvate transport in working guinea-pig heart. Work-related vs. carrier-mediated control of pyruvate oxidation. Biochim. Biophys. Acta, 1151, 223–236. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
