

A split motor domain in a cytoplasmic dynein
- PMID: 11566874
- PMCID: PMC125636
- DOI: 10.1093/emboj/20.18.5091
A split motor domain in a cytoplasmic dynein
Abstract
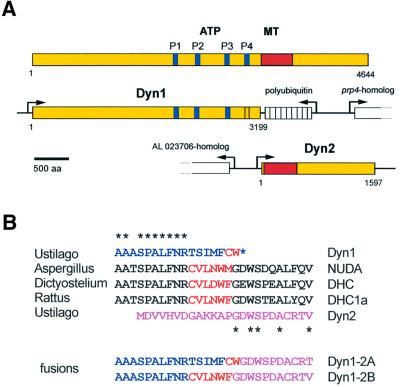
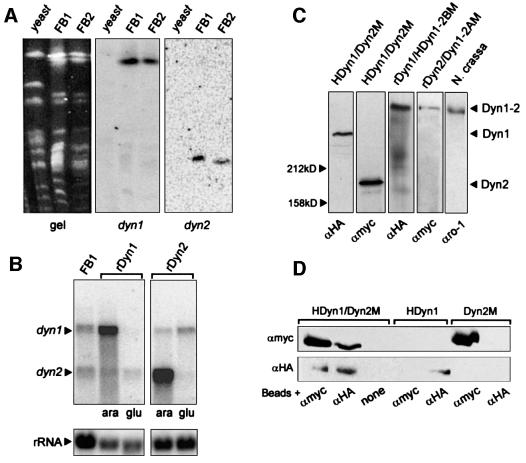
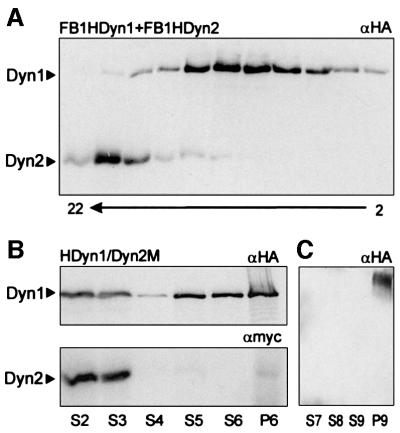
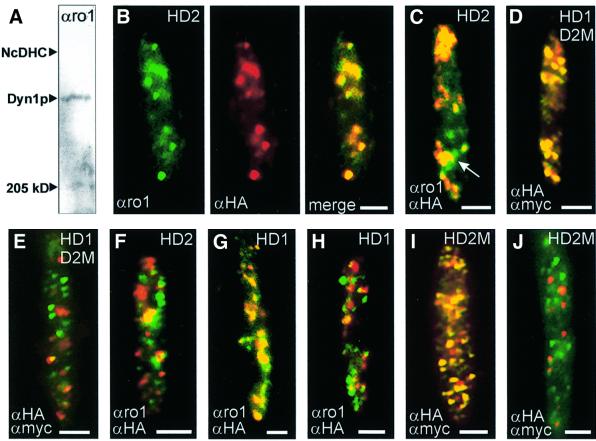
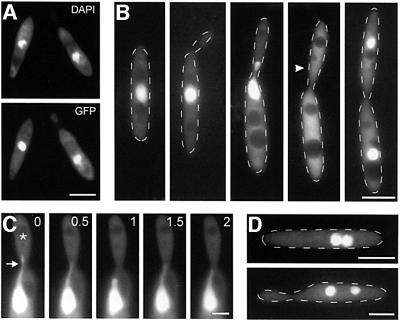
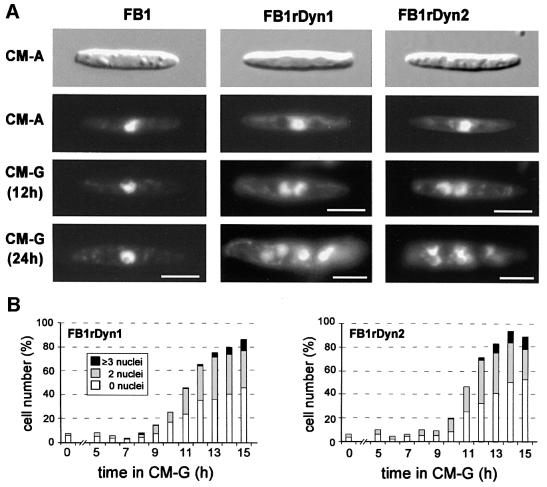
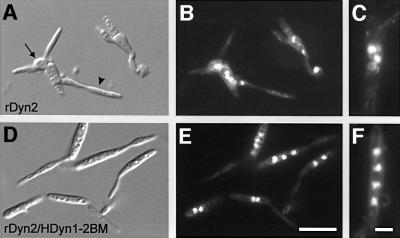
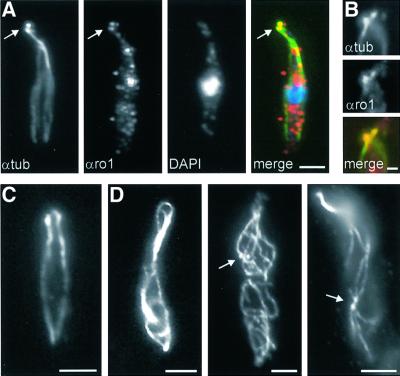
The heavy chain of dynein forms a globular motor domain that tightly couples the ATP-cleavage region and the microtubule-binding site to transform chemical energy into motion along the cytoskeleton. Here we show that, in the fungus Ustilago maydis, two genes, dyn1 and dyn2, encode the dynein heavy chain. The putative ATPase region is provided by dyn1, while dyn2 includes the predicted microtubule-binding site. Both genes are located on different chromosomes, are transcribed into independent mRNAs and are translated into separate polypeptides. Both Dyn1 and Dyn2 co-immunoprecipitated and co-localized within growing cells, and Dyn1-Dyn2 fusion proteins partially rescued mutant phenotypes, suggesting that both polypeptides interact to form a complex. In cell extracts the Dyn1-Dyn2 complex dissociated, and microtubule affinity purification indicated that Dyn1 or associated polypeptides bind microtubules independently of Dyn2. Both Dyn1 and Dyn2 were essential for cell survival, and conditional mutants revealed a common role in nuclear migration, cell morphogenesis and microtubule organization, indicating that the Dyn1-Dyn2 complex serves multiple cellular functions.
Figures
Similar articles
-
The yeast dynein Dyn2-Pac11 complex is a dynein dimerization/processivity factor: structural and single-molecule characterization.Mol Biol Cell. 2013 Aug;24(15):2362-77. doi: 10.1091/mbc.E13-03-0166. Epub 2013 Jun 12. Mol Biol Cell. 2013. PMID: 23761070 Free PMC article.
-
Dynein supports motility of endoplasmic reticulum in the fungus Ustilago maydis.Mol Biol Cell. 2002 Mar;13(3):965-77. doi: 10.1091/mbc.01-10-0475. Mol Biol Cell. 2002. PMID: 11907275 Free PMC article.
-
A balance of KIF1A-like kinesin and dynein organizes early endosomes in the fungus Ustilago maydis.EMBO J. 2002 Jun 17;21(12):2946-57. doi: 10.1093/emboj/cdf296. EMBO J. 2002. PMID: 12065408 Free PMC article.
-
Nuclear migration and positioning in filamentous fungi.Fungal Genet Biol. 2004 Apr;41(4):411-9. doi: 10.1016/j.fgb.2003.11.010. Fungal Genet Biol. 2004. PMID: 14998524 Review.
-
The role of microtubules in cellular organization and endocytosis in the plant pathogen Ustilago maydis.J Microsc. 2004 May;214(Pt 2):114-23. doi: 10.1111/j.0022-2720.2004.01319.x. J Microsc. 2004. PMID: 15102060 Review.
Cited by
-
Motor-driven motility of fungal nuclear pores organizes chromosomes and fosters nucleocytoplasmic transport.J Cell Biol. 2012 Aug 6;198(3):343-55. doi: 10.1083/jcb.201201087. Epub 2012 Jul 30. J Cell Biol. 2012. PMID: 22851316 Free PMC article.
-
Nuclear movement in fungi.Semin Cell Dev Biol. 2018 Oct;82:3-16. doi: 10.1016/j.semcdb.2017.10.024. Epub 2017 Dec 11. Semin Cell Dev Biol. 2018. PMID: 29241689 Free PMC article. Review.
-
The key protein of endosomal mRNP transport Rrm4 binds translational landmark sites of cargo mRNAs.EMBO Rep. 2019 Jan;20(1):e46588. doi: 10.15252/embr.201846588. Epub 2018 Dec 14. EMBO Rep. 2019. PMID: 30552148 Free PMC article.
-
Controlled and stochastic retention concentrates dynein at microtubule ends to keep endosomes on track.EMBO J. 2011 Feb 16;30(4):652-64. doi: 10.1038/emboj.2010.360. Epub 2011 Jan 28. EMBO J. 2011. PMID: 21278707 Free PMC article.
-
New insights into the mechanism of dynein motor regulation by lissencephaly-1.Elife. 2020 Jul 21;9:e59737. doi: 10.7554/eLife.59737. Elife. 2020. PMID: 32692650 Free PMC article.
References
-
- Asai D.J and Koonce,M.P. (2001) The dynein heavy chain: structure, mechanics and evolution. Trends Cell Biol., 11, 196–202. - PubMed
-
- Banuett F. (1995) Genetics of Ustilago maydis, a fungal pathogen that induces tumors in maize. Annu. Rev. Genet., 29, 179–208. - PubMed
-
- Bölker M., Bohnert,H.U., Braun,K.H., Görl,J. and Kahmann,R. (1995) Tagging pathogenicity genes in Ustilago maydis by restriction-enzyme mediated integration (REMI). Mol. Gen. Genet., 248, 547–552. - PubMed
-
- Bottin A., Kämper,J. and Kahmann,R. (1996) Isolation of a carbon source-regulated gene from Ustilago maydis. Mol. Gen. Genet., 253, 342–352. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
