

Nonspecific adherence and fibril biogenesis by Actinobacillus actinomycetemcomitans: TadA protein is an ATPase
- PMID: 11566992
- PMCID: PMC99671
- DOI: 10.1128/JB.183.20.5927-5936.2001
Nonspecific adherence and fibril biogenesis by Actinobacillus actinomycetemcomitans: TadA protein is an ATPase
Abstract
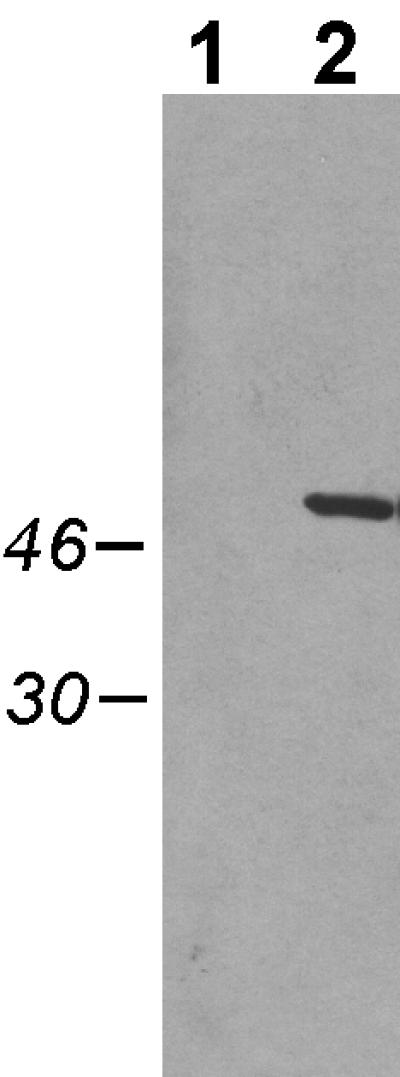
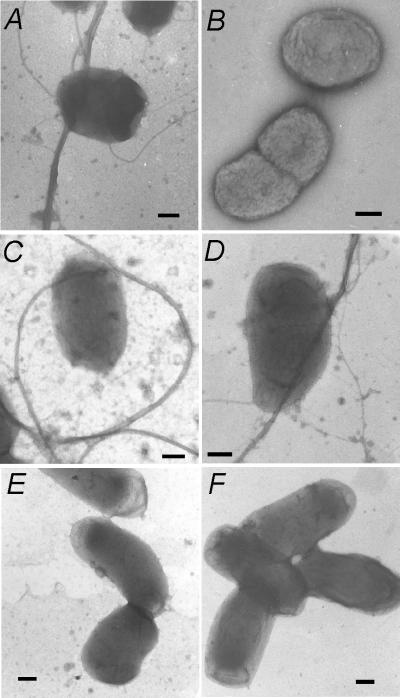
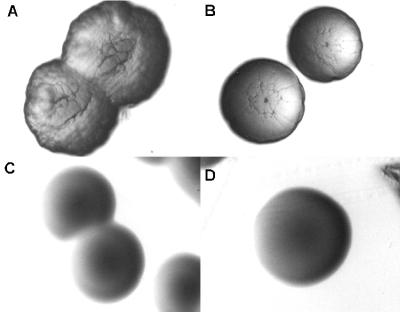
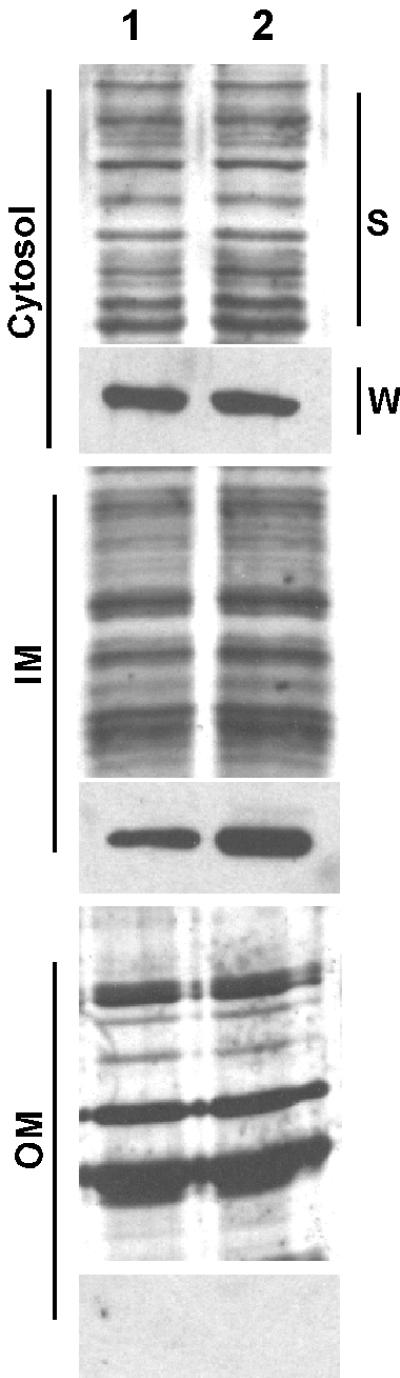
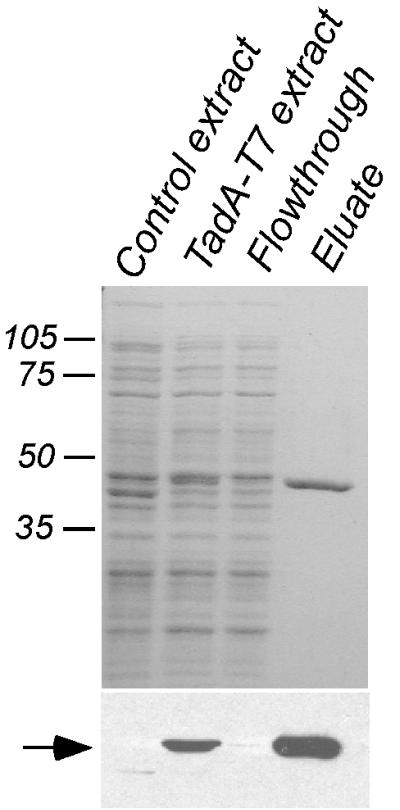
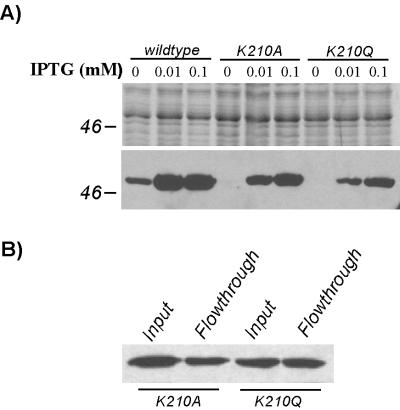
Cells of Actinobacillus actinomycetemcomitans, a gram-negative pathogen responsible for an aggressive form of juvenile periodontitis, form tenaciously adherent biofilms on solid surfaces. The bacteria produce long fibrils of bundled pili, which are required for adherence. Mutations in flp-1, which encodes the major subunit of the pili, or any of seven downstream tad genes (tadABCDEFG) cause defects in fibril production, autoaggregation, and tenacious adherence. We proposed that the tad genes specify part of a novel secretion system for the assembly and transport of Flp pili. The predicted amino acid sequence of TadA (426 amino acids, 47,140 Da) contains motifs for nucleotide binding and hydrolysis common among secretion NTP hydrolase (NTPase) proteins. In addition, the tadA gene is the first representative of a distinct subfamily of potential type IV secretion NTPase genes. Here we report studies on the function of TadA. The tadA gene was altered to express a modified version of TadA that has the 11-residue epitope (T7-TAG) fused to its C terminus. The TadA-T7 protein was indistinguishable from the wild type in its ability to complement the fibril and adherence defects of A. actinomycetemcomitans tadA mutants. Although TadA is not predicted to have a transmembrane domain, the protein was localized to the inner membrane and cytoplasmic fractions of A. actinomycetemcomitans cells, indicating a possible peripheral association with the inner membrane. TadA-T7 was purified and found to hydrolyze ATP in vitro. The ATPase activity is stimulated by Triton X-100, with maximal stimulation at the critical micellar concentration. TadA-T7 forms multimers that are stable during sodium dodecyl sulfate-polyacrylamide gel electrophoresis in nonreducing conditions, and electron microscopy revealed that TadA-T7 can form structures closely resembling the hexameric rings of other type IV secretion NTPases. Site-directed mutagenesis was used to substitute Ala and Gln residues for the conserved Lys residue of the Walker A box for nucleotide binding. Both mutants were found to be defective in their ability to complement tadA mutants. We suggest that the ATPase activity of TadA is required to energize the assembly or secretion of Flp pili for tight adherence of A. actinomycetemcomitans.
Figures



References
-
- Aepfelbacher M, Zumbihl R, Ruckdeschel K, Jacobi C A, Barz C, Heesemann J. The tranquilizing injection of Yersinia proteins: a pathogen's strategy to resist host defense. Biol Chem. 1999;380:795–802. - PubMed
-
- Ausubel F M. Short protocols in molecular biology: a compendium of methods from Current Protocols in Molecular Biology. New York, N.Y: Greene Publishing Associates and Wiley-Interscience; 1989.
-
- Binet R, Letoffe S, Ghigo J M, Delepelaire P, Wandersman C. Protein secretion by gram-negative bacterial ABC exporters—a review. Gene. 1997;192:7–11. - PubMed
-
- Burns D L. Biochemistry of type IV secretion. Curr Opin Microbiol. 1999;2:25–29. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
