

Neurons lacking huntingtin differentially colonize brain and survive in chimeric mice
- PMID: 11567051
- PMCID: PMC6762912
- DOI: 10.1523/JNEUROSCI.21-19-07608.2001
Neurons lacking huntingtin differentially colonize brain and survive in chimeric mice
Abstract
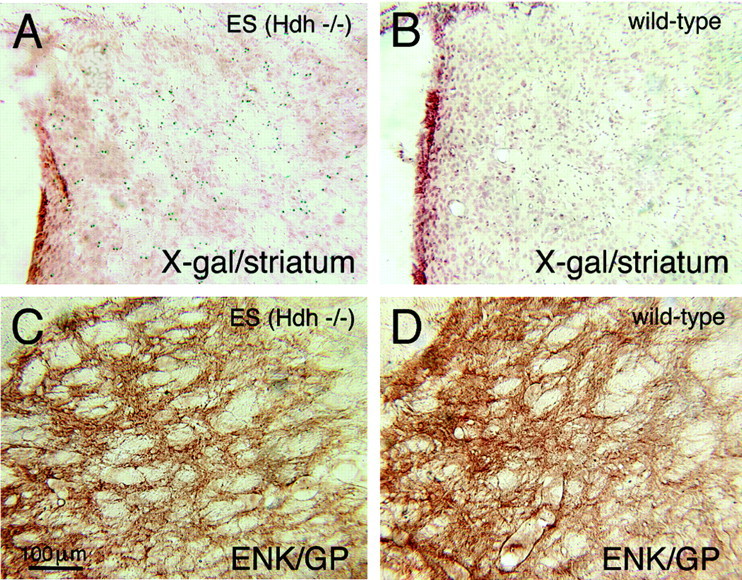
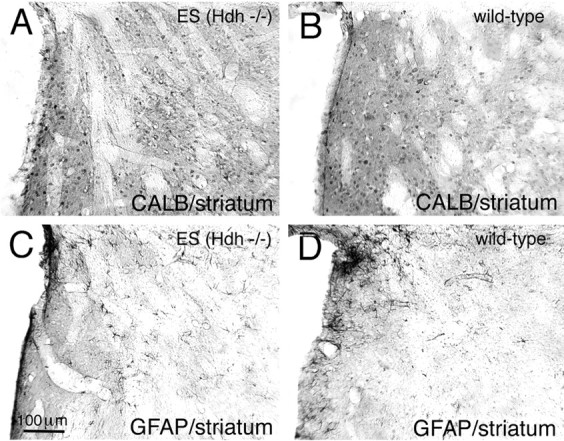
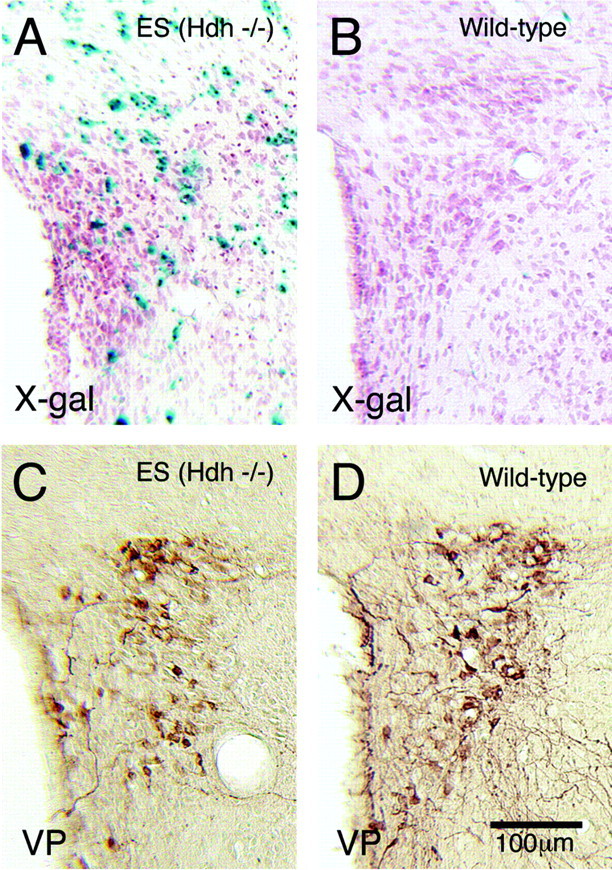
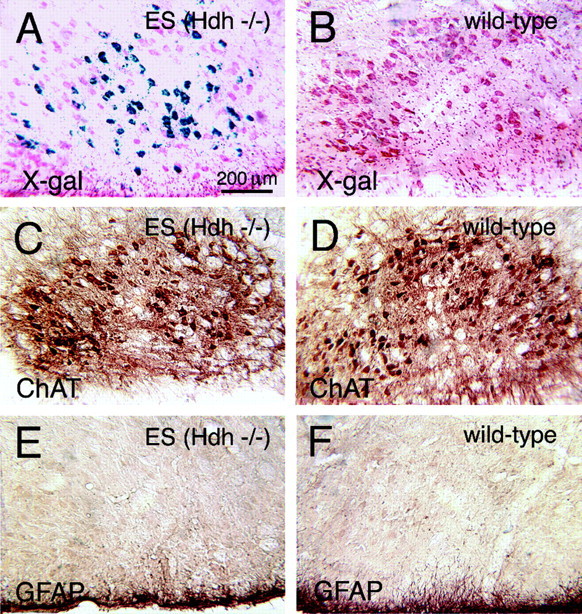
To determine whether neurons lacking huntingtin can participate in development and survive in postnatal brain, we used two approaches in an effort to create mice consisting of wild-type cells and cells without huntingtin. In one approach, chimeras were created by aggregating the 4-8 cell embryos from matings of Hdh (+/-) mice with wild-type 4-8 cell embryos. No chimeric offspring that possessed homozygous Hdh (-/-) cells were obtained thereby, although statistical considerations suggest that such chimeras should have been created. By contrast, Hdh (-/-) ES cells injected into blastocysts yielded offspring that were born and in adulthood were found to have Hdh (-/-) neurons throughout brain. The Hdh (-/-) cells were, however, 5-10 times more common in hypothalamus, midbrain, and hindbrain than in telencephalon and thalamus. Chimeric animals tended to be smaller than wild-type littermates, and chimeric mice rich in Hdh (-/-) cells tended to show motor abnormalities. Nonetheless, no brain malformations or pathologies were evident. The apparent failure of aggregation chimeras possessing Hdh (-/-) cells to survive to birth is likely attributable to the previously demonstrated critical role of huntingtin in extraembryonic membranes. That Hdh (-/-) cells in chimeric mice created by blastocyst injection are under-represented in adult telencephalon and thalamus implies a role for huntingtin in the development of these regions, whereas the neurological dysfunction in brains enriched in Hdh (-/-) cells suggests a role for huntingtin in adult brain. Nonetheless, the lengthy survival of Hdh (-/-) cells in adult chimeric mice indicates that individual neurons in many brain regions do not require huntingtin to participate in normal brain development and to survive.
Figures



References
-
- Albin RL, Tagle DA. Genetics and molecular biology of Huntington's disease. Trends Neurosci. 1995;18:11–14. - PubMed
-
- Albin RL, Reiner A, Anderson KD, Penney JB, Young AB. Striatal and nigral neuron subpopulations in rigid Huntington's disease: implications for the functional anatomy of chorea and rigidity-akinesia. Ann Neurol. 1990a;27:357–365. - PubMed
-
- Albin RL, Young AB, Penney JB, Handelin B, Balfour R, Anderson KD, Markel DS, Tourtellotte WW, Reiner A. Abnormalities of striatal projection neurons and N-methyl-d-aspartate receptors in presymptomatic Huntington's disease. N Engl J Med. 1990b;332:1293–1298. - PubMed
-
- Albin RL, Reiner A, Anderson KD, Dure LS, IV, Handelin B, Balfour R, Whetsell WO, Jr, Penney JB, Young AB. Preferential loss of striato-external pallidal projection neurons in presymptomatic Huntington's disease. Ann Neurol. 1992;31:425–430. - PubMed
-
- Ambrose CM, Duyao MP, Barnes G, Bates GP, Lin CS, Srinidhi J, Baxendale S, Hummerich H, Lehrach H, Atherr M, Wasmuth J, Buckler A, Church D, Housman D, Berks M, Micklem G, Durbin R, Dodge A, Read A, Gusella J, MacDonald ME. Evidence against simple inactivation due to an expanded CAG repeat. Somat Cell Mol Genet. 1994;20:27–38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases