

Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4
- PMID: 11574472
- PMCID: PMC125652
- DOI: 10.1093/emboj/20.19.5400
Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4
Abstract
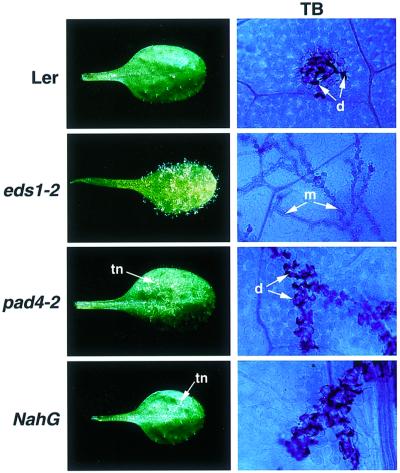
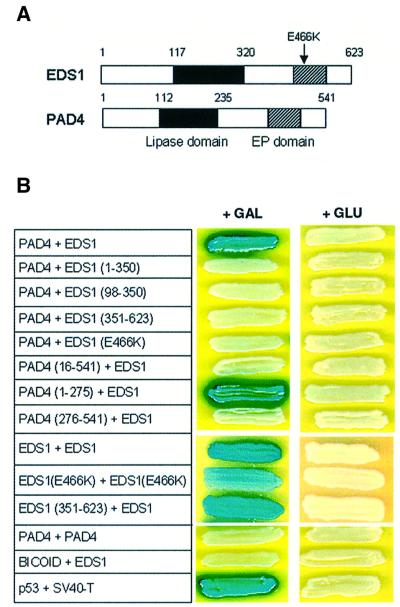
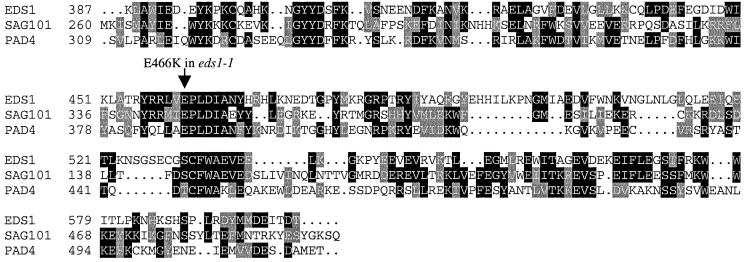
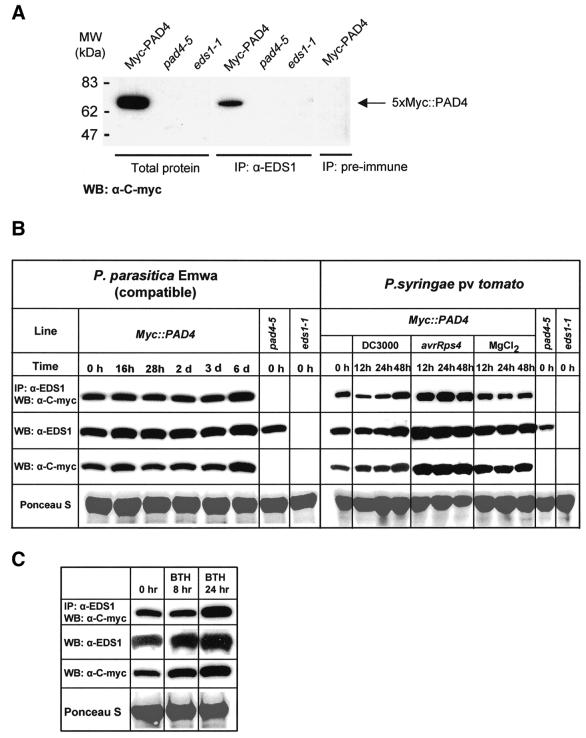
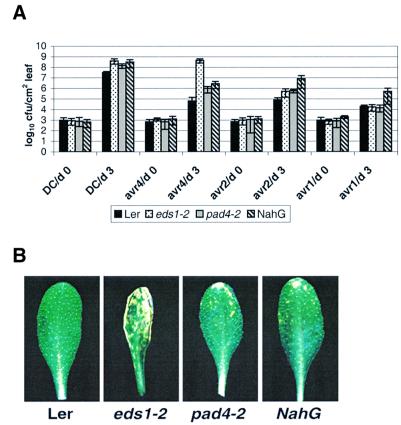
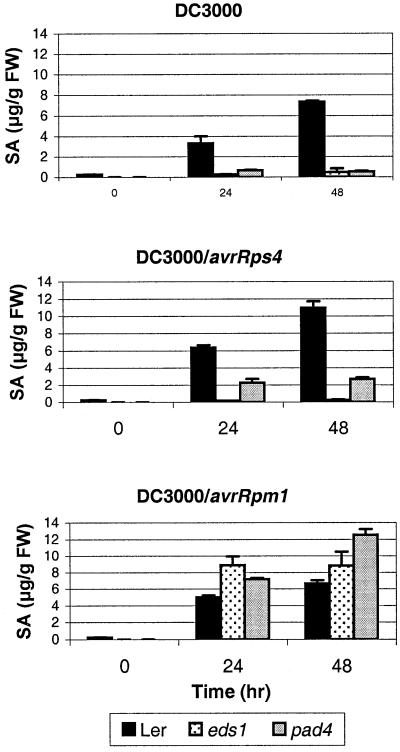
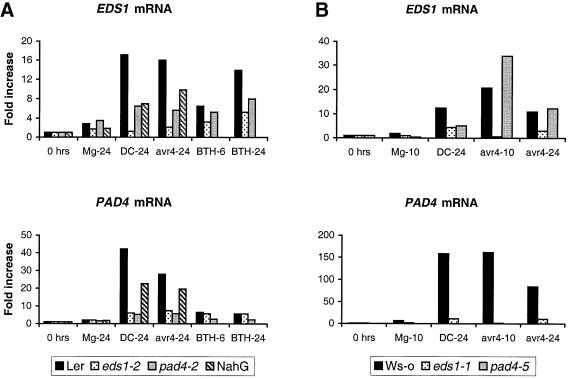
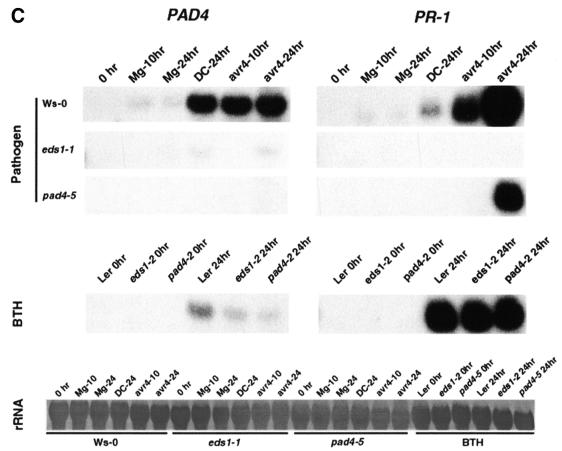
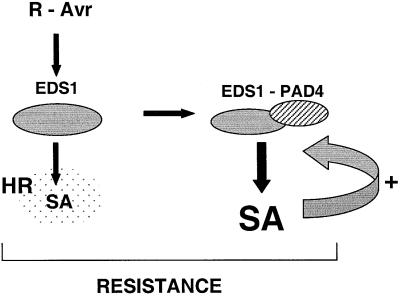
The Arabidopsis EDS1 and PAD4 genes encode lipase-like proteins that function in resistance (R) gene-mediated and basal plant disease resistance. Phenotypic analysis of eds1 and pad4 null mutants shows that EDS1 and PAD4 are required for resistance conditioned by the same spectrum of R genes but fulfil distinct roles within the defence pathway. EDS1 is essential for elaboration of the plant hypersensitive response, whereas EDS1 and PAD4 are both required for accumulation of the plant defence-potentiating molecule, salicylic acid. EDS1 is necessary for pathogen-induced PAD4 mRNA accumulation, whereas mutations in PAD4 or depletion of salicylic acid only partially compromise EDS1 expression. Yeast two-hybrid analysis reveals that EDS1 can dimerize and interact with PAD4. However, EDS1 dimerization is mediated by different domains to those involved in EDS1-PAD4 association. Co-immunoprecipitation experiments show that EDS1 and PAD4 proteins interact in healthy and pathogen-challenged plant cells. We propose two functions for EDS1. The first is required early in plant defence, independently of PAD4. The second recruits PAD4 in the amplification of defences, possibly by direct EDS1-PAD4 association.
Figures
References
-
- An Y.Q., McDowell,J.M., Huang,S.R., McKinney,E.C., Chambliss,S. and Meagher,R.B. (1996) Strong, constitutive expression of the Arabidopsis ACT2/ACT8 actin subclass in vegetative tissues. Plant J., 10, 107–121. - PubMed
-
- Ausubel F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (1996) Current Protocols in Molecular Biology. John Wiley & Sons, New York, NY.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
