

The proto-oncoprotein Brx activates estrogen receptor beta by a p38 mitogen-activated protein kinase pathway
- PMID: 11579095
- PMCID: PMC4152864
- DOI: 10.1074/jbc.M106927200
The proto-oncoprotein Brx activates estrogen receptor beta by a p38 mitogen-activated protein kinase pathway
Abstract
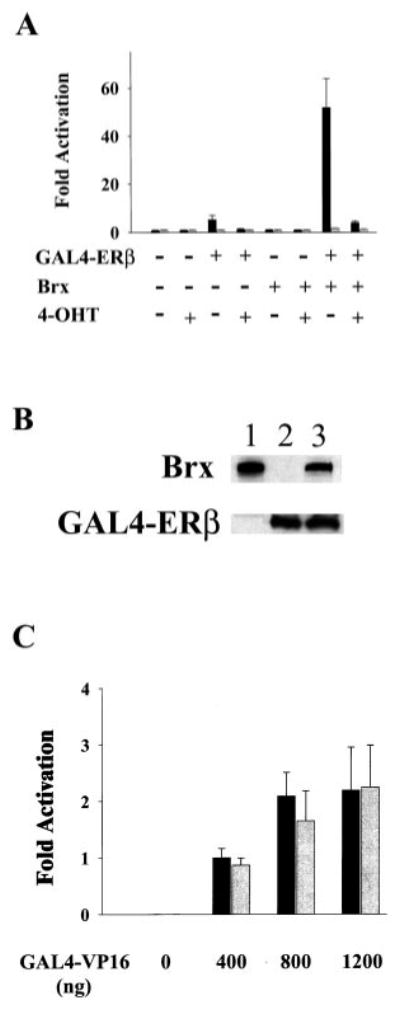
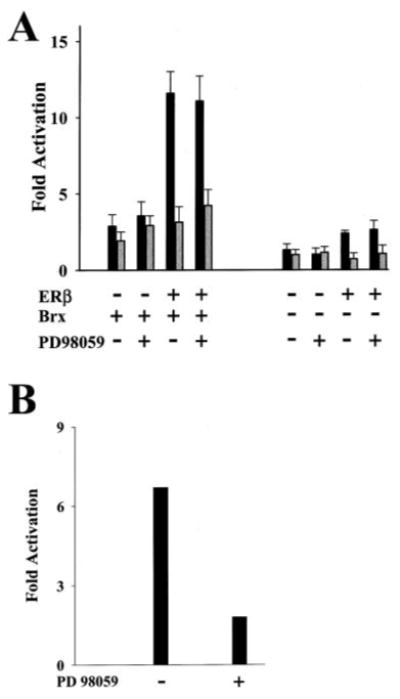
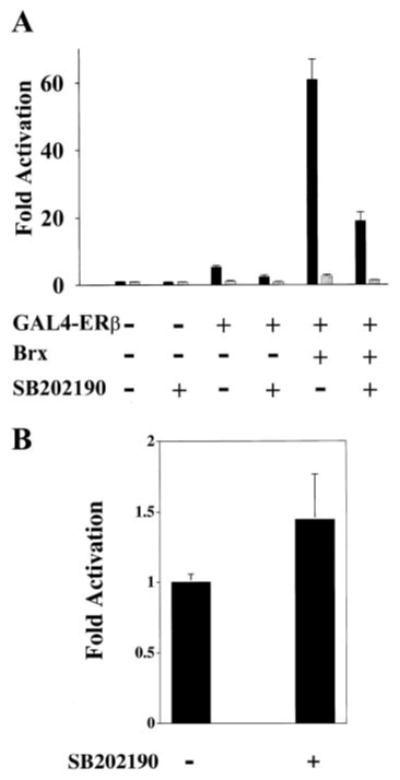
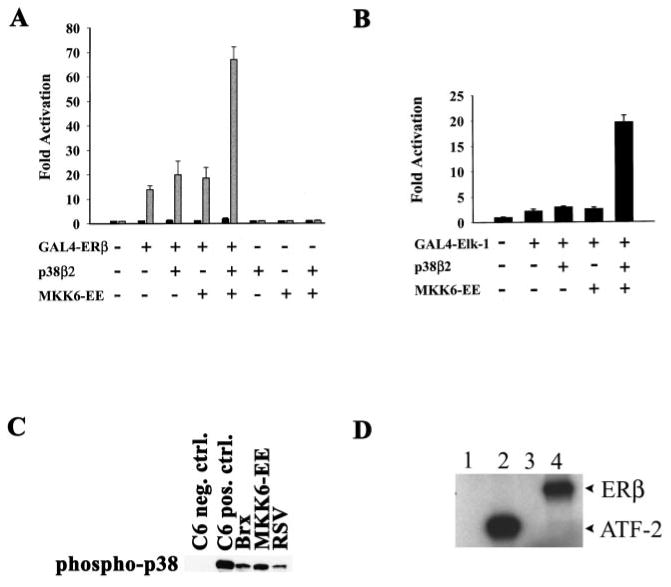
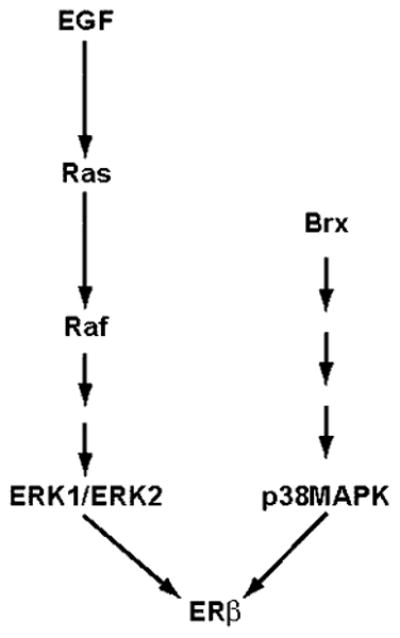
The estrogen receptors (ERs) are ligand-inducible transcription factors that play key roles in the control of growth and differentiation in reproductive tissues. We showed that the novel Dbl family proto-oncoprotein Brx enhances ligand-dependent activity of ERalpha via a Cdc42-dependent pathway. Brx also significantly enhances ligand-dependent activity of ERbeta. This enhancement is not affected by inhibition of p44/42 mitogen-activated protein kinase (MAPK) activation by PD98059. However, addition of the p38 MAPK inhibitor SB202190 abrogates the enhancement of ERbeta activity by Brx, showing that p38 MAPK activity is required for the enhancement of ERbeta function by Brx. In COS-7 cells, transfection of Brx leads to activation of endogenous p38 MAPK activity. Co-expression of the beta2 isoform of human p38 MAPK and a constitutively active form of the p38 MAPK kinase MKK6 (MKK6-EE) synergistically augments ligand-dependent activity of ERbeta. Our findings suggest that p38 MAPKs may be important regulators of ERbeta activity.
Figures
References
-
- Mosselman S, Polman J, Dijkema R. FEBS Lett. 1996;392:49–53. - PubMed
-
- Tsai M, O'Malley BW. Annu Rev Biochem. 1994;63:451–486. - PubMed
-
- Tora L, White J, Brou C, Tasset D, Webster N, Scheer E, Chambon P. Cell. 1989;59:477–487. - PubMed
-
- O'Malley BW, Schrader WT, Mani S, Smith C, Weigel NL, Conneely OM, Clark JH. Recent Prog Horm Res. 1995;50:333–347. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
